RNR -polimerazė yra fermentas, sintetinantis RNR molekules. Siaurąja prasme RNR polimerazė paprastai vadinama nuo DNR priklausomomis RNR polimerazėmis, kurios sintezuoja RNR molekules ant DNR šablono, tai yra, atlieka transkripciją. RNR polimerazės klasės fermentai yra labai svarbūs ląstelės funkcionavimui, todėl jų yra visuose organizmuose ir daugelyje virusų. Chemiškai RNR polimerazės yra nukleotidilo transferazės, kurios polimerizuoja ribonukleotidus 3 colių RNR grandinės gale.
Promotorius yra DNR nukleotidų seka, kurią RNR polimerazė atpažįsta kaip specifinės arba reikšmingos transkripcijos pradžios tašką. Prokariotuose promotorius apima daugybę motyvų, kurie yra svarbūs jį atpažįstant RNR polimeraze, ypač vadinamąsias -10 ir -35 sekas. Promotorius yra asimetriškas, o tai leidžia RNR polimerazei pradėti transkripciją teisinga kryptimi ir nurodo, kuri iš dviejų DNR grandinių bus RNR sintezės šablonas.
Protoriaus sritis operone gali iš dalies persidengti arba visai nesutapti su cistrono (geno) operatoriaus sritimi.
Promotorius, po kuriuo yra RNR koduojanti DNR sritis, vaidina lemiamą vaidmenį šio geno ekspresijos intensyvumui kiekviename specifiniame ląstelių tipe. Promotorių aktyvavimą lemia tai, kad kiekviename ląstelių tipe yra skirtingas transkripcijos faktorių rinkinys.
Terminatorius yra DNR nukleotidų seka, kurią RNR polimerazė atpažįsta kaip signalą sustabdyti RNR molekulės sintezę ir transkripcijos komplekso disociaciją.
Dažnai šios sekos baigiasi timino nukleotidų grandine (transkriptuose - uridinas), prieš kurią yra sritis, kurioje yra vidinės, viena kitą papildančios sekos priešingomis kryptimis („plaukų segtuko“ struktūros). Plaukų segtuko sritis yra praturtinta GC poromis, todėl ši struktūra yra stabilesnė.
78. Transkripcijos proceso skirtumai skirtingose ląstelėse. Mozaikinė genų struktūra. Heterobranduolinės RNR sintezė, jos pavertimas pasiuntinėmis RNR. Sujungimas. Apdorojimas.
Kai kurie virusai (pvz., ŽIV, sukeliantis AIDS) turi galimybę perrašyti RNR į DNR. ŽIV turi RNR genomą, kuris yra integruotas į DNR. Dėl to viruso DNR gali būti sujungta su ląstelės šeimininkės genomu. Pagrindinis fermentas, atsakingas už DNR sintezę iš RNR, vadinamas atvirkštiniu. Viena iš reversetazės funkcijų yra sukurti komplementarią DNR (cDNR) iš viruso genomo. Susijęs fermentas ribonukleazė H skaldo RNR, o reverseazė sintetina cDNR iš DNR dvigubos spiralės. cDNR integrazės dėka integruojama į šeimininko ląstelės genomą. Rezultatas yra viruso baltymų sintezė ląstelėje-šeimininkėje, kuri sudaro naujus virusus. ŽIV atveju taip pat užprogramuota T-limfocitų apoptozė (ląstelių mirtis). Kitais atvejais ląstelė gali likti virusų platintoja.
Kai kuriose eukariotinėse ląstelėse yra fermento telomerazės, kuri taip pat pasižymi atvirkštinės transkripcijos aktyvumu. Su jo pagalba sintezuojamos pasikartojančios sekos DNR. Telomerazė dažnai aktyvuojama vėžio ląstelėse, kad neribotą laiką dubliuotų genomą, neprarandant baltymą koduojančios DNR sekos.
Išskirtinis daugelio eukariotų genų struktūros bruožas yra mozaikinė (nepertraukiama) semantinės genų dalies struktūra.
Šis skirtumas yra susijęs su semantinių regionų, pernešančių informaciją apie aminorūgščių seką baltyme, kaitaliojimu – egzonais ir nekoduojančių sekų, kurios yra žymiai ilgesnės už egzonus – intronų, kaita. Dėl to bendras geno ilgis yra maždaug 5–7 kartus ilgesnis nei būtų galima tikėtis.
Geno pradžioje (prieš jo semantinę dalį) yra sritys, kurios užtikrina tinkamą geno reguliavimą. Jie skirstomi į dvi grupes: nespecifinius (visiems genams vienodi) ir specifinius (būdingus tik tam tikram genui). Nespecifinės reguliavimo sritys eukariotuose vadinamos „TATA-BOX“. Jie pakaitomis vartoja timiną ir adeniną. Ši sritis yra 30 nukleotidų kairėje nuo geno skaitymo pradžios. Nustatyta, kad RNR polimerazė remiasi į DNR taip, kad jos atpažinimo dalis uždaro TATA BOX, o aktyvus RNR polimerazės centras atsiranda virš pirmojo nuskaityto nukleotido. Po to seka promotoriaus sritis, susidedanti iš atpažinimo vietos, surišimo vietos ir transkripcijos iniciacijos vietos (A arba D). Nukleotidų derinys promotoriuje yra toks, kad nuskaitymo rėmelį nustačius neteisingai, gaunama stopkodonams būdinga seka ir transkripcija sustoja. Po promotoriaus srities seka palindromas („apversti“) arba apverstas kartojimas. Tai dvigrandė DNR seka, kuri abiem kryptimis skaitoma vienodai („kazokas“ – iš dešinės į kairę ir iš kairės į dešinę skaito tą patį). Kartais tokia seka vadinama „dvigubai simetriška sritimi“, kur simetrijos ašis yra centrinis taškas, kurio atžvilgiu seka išlieka ta pati. Svarbi palindromų savybė yra galimybė suformuoti plaukų segtukų struktūras RNR arba kryžmines struktūras DNR. Jei skaitymo rėmelis sumontuotas neteisingai, DNR palindromas virsta „kryžiu“, todėl tolesnis fermento progresas tampa neįmanomas. Kartais palindromai gali būti geno gale po jo semantinės dalies ir, taip pat, sudarydami „kryželį“, sustabdys mRNR sintezę. Semantinė geno dalis, kaip rašėme aukščiau, susideda iš kintančių intronų-egzonų sekų. Struktūrinėje geno dalyje yra promotorius ir palindromas.
Pastaruoju metu buvo nustatyti specifinio genų reguliavimo intarpai – stiprintuvai. Jie yra prieš geną šimtų ir tūkstančių nukleotidų porų atstumu. Eukariotai turi specialius reguliuojančius baltymus, kurie atpažįsta stipriklį ir prie jo prisijungia. Taip suaktyvinamas genas. Genomą formuoja ne tik branduoliniai, bet ir citoplazminiai genai. Genome genai atlieka skirtingas funkcijas. Priklausomai nuo jų atliekamų funkcijų, genai yra skirstomi į privalomus genus, kurie koduoja gyvybines funkcijas visų tipų ląstelėse (kartais vadinami „namų priežiūros“ genais), struktūrinius, reguliavimo genus ir skyrybos genus. Nors prokariotinės (bakterinės ir archeologinės) mRNR, išskyrus retas išimtis, yra iš karto paruoštos vertimui ir joms nereikia specialaus apdorojimo, eukariotinės pre-mRNR intensyviai modifikuojamos. Taigi, kartu su transkripcija, „redagavimas“ (splaisavimas) vyksta jau susintetintoje mRNR dalyje. Splaisingo proceso metu iš pre-mRNR pašalinamos baltymų nekoduojančios sekos – intronai, į 5" molekulės galą pridedamas specialus modifikuotas nukleotidas (dangtelis), o keli adeninai, vadinamoji poliadenino uodega, pridedami prie 3 colių galo. Dangtelis atpažįstamas pagal iniciacijos faktorius, baltymus, atsakingus už prisijungimą prie ribosomos mRNR, o poliadenino uodega jungiasi prie specialaus baltymo PAB. Paprastai šie eukariotų mRNR pokyčiai po transkripcijos vadinami „mRNR apdorojimu“. Poliadenilinimas yra būtinas daugumos mRNR transportavimui į citoplazmą ir apsaugo mRNR molekules nuo greito skilimo (pailgina jų pusinės eliminacijos periodą). mRNR molekulės, neturinčios poli-A srities (pavyzdžiui, virusinės), ribonukleazės greitai sunaikinamos eukariotinių ląstelių citoplazmoje.
Sujungimas (iš anglų kalbos splice - sujungti ar suklijuoti ko nors galus) yra tam tikrų nukleotidų sekų išpjovimas iš RNR molekulių ir sekų, kurios RNR apdorojimo metu lieka „subrendusioje“ molekulėje, sujungimas.Šis procesas dažniausiai vyksta eukariotuose bręstant pasiuntinei RNR (mRNR), kurio metu per biochemines reakcijas, kuriose dalyvauja RNR ir baltymai, pašalinamos baltymo nekoduojančios mRNR dalys (intronai), o sekcijos, koduojančios amino. rūgščių seka – egzonai yra sujungti vienas su kitu. Taigi nesubrendusi pre-mRNR paverčiama subrendusia iRNR, iš kurios skaitomi (verčiami) ląstelės baltymai. Dauguma prokariotų baltymus koduojančių genų neturi intronų, todėl pre-mRNR susijungimas juose yra retas. Pernešimo RNR (tRNR) ir kitų nekoduojančių RNR susijungimas taip pat vyksta eukariotų, bakterijų ir archajų atstovuose.
RNR apdorojimas
Tarp transkripcijos ir transliacijos iRNR molekulėje vyksta eilė nuoseklių pokyčių, kurie užtikrina funkcionuojančios matricos subrendimą polipeptidinės grandinės sintezei. Prie 5΄ galo pritvirtintas dangtelis, o prie 3΄ galo pritvirtintas poli-A uodega, o tai padidina mRNR tarnavimo laiką. Atsiradus apdorojimui eukariotų ląstelėje, atsirado galimybė sujungti genų egzonus, kad būtų gauta didesnė baltymų, užkoduotų viena DNR nukleotidų seka, įvairovė – alternatyvus sujungimas.
Norint atlikti teisingą transkripciją, reikalingi dviejų tipų reguliavimo elementai. Pirmojo tipo reguliavimo elementai vadinami cis-reguliatoriai. Tai yra specifinės DNR sekos tam tikroje chromosomoje. Cis -reguliatoriai veikia tik šalia esančius genus. Antrasis tipas vadinamas transreguliatoriai. Tai yra tirpios molekulės (įskaitant baltymus ir RNR), kurias gamina vienas genas ir kurios sąveikauja su kitais genais toje pačioje chromosomoje arba kitose chromosomose. Jei pereisime prie genų indukcijos lac - operonasE., coli Tai yra tirpios molekulės (įskaitant baltymus ir RNR), kurias gamina vienas genas ir kurios sąveikauja su kitais genais toje pačioje chromosomoje arba kitose chromosomose. Jei pereisime prie genų indukcijos tada galime prisiminti, kad represoriaus genas gamina represoriaus baltymą, kuris sąveikauja su genų operatorių seka - operonas.Šiuo atveju operatorius yra Tai yra tirpios molekulės (įskaitant baltymus ir RNR), kurias gamina vienas genas ir kurios sąveikauja su kitais genais toje pačioje chromosomoje arba kitose chromosomose. Jei pereisime prie genų indukcijos- cis – reguliavimo elementas, nes jis tik kontroliuoja savo chromosomos operonas. - operonas.(Kitoje chromosomoje esanti mutantinė operatoriaus seka gali prijungti arba neprijungti represoriaus baltymo.) Represoriaus baltymas, priešingai, yra
transas - operonas.- reguliatorius. nes jį gamina viena chromosoma ir jungiasi prie paprastai yra prieš pat vietą, kurioje ji prasideda
Gilbert s. Raidos biologija: 3 tomai T. 2: Vert. Iš anglų kalbos – M.: Mir, 1994. – 235 p.
142 _______________ 12 SKYRIUS _____________________________________________________________________________
|
Ryžiai. 12.5. Diferencinio genų reguliavimo schema in E. E.; parodyta - operonas.- Ir – reguliavimo elementas, nes jis tik kontroliuoja-reguliavimo elementai. Laukinio tipo ląstelėse indukuojama būsena pasižymi tuo, kad β-galaktozidazės RNR nėra transkribuojama tol, kol nėra laktozės. Trūkstant laktozės, represoriaus baltymo (R), koduojamo geno i, prisijungia prie operatoriaus svetainės ( O), tokiu būdu slopinant RNR polimerazės transkripciją iš promotoriaus ( p). E. Jei yra laktozės, ji jungiasi su represoriaus baltymu, todėl represorius negali prisijungti prie DNR ir transkripcija tęsiasi. Šio represoriaus tirpumas buvo parodytas atliekant eksperimentus su mutantais. Coli i – Kai haploidinės bakterijų ląstelės, turinčios geną i, tampa iš dalies diploidinis su genu i + laukinis tipas ( – reguliavimo elementas, nes jis tik kontroliuoja), sintetinamas laukinio tipo represorius, galintis padaryti originalų ß-galaktozidazės geną indukuojamą. - operonas.Šis represoriaus baltymas yra |
|
|
|
-reguliacinis elementas. Promotorių ir operatorių sekos yra |

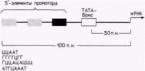
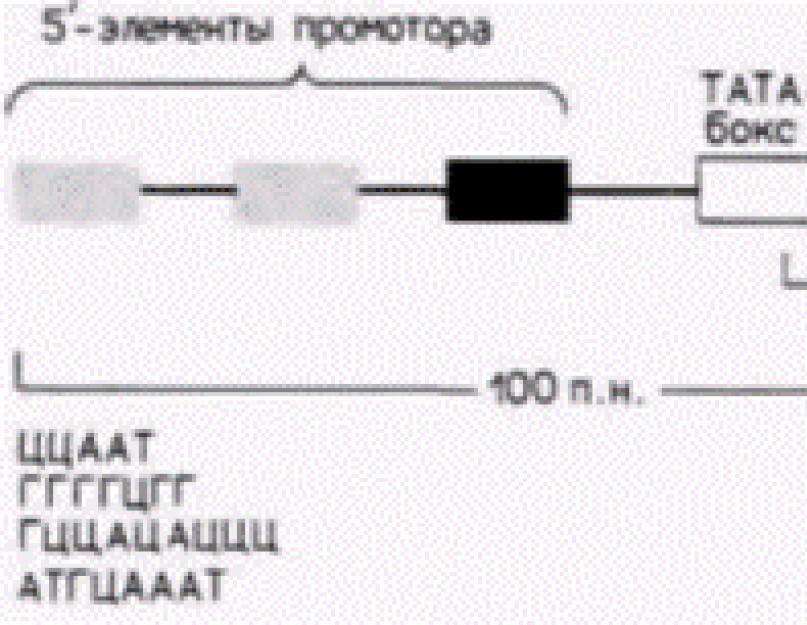
-reguliavimo elementai. - operonas. Ryžiai. 12.6. Tipiškas eukariotinį baltymą koduojančio geno promotorius. Pateiktame gene yra TATA dėžutė ir trys 5" promotoriaus elementai. Tokių 5' elementų pavyzdžiai pateikti apatinėje paveikslo dalyje. (After Maniatis ir kt., 1987).
transkripcija ir yra maždaug 100 bazinių porų ilgio. Protoriaus sritis reikalinga RNR polimerazės II surišimui ir tiksliam transkripcijos inicijavimui. Stipriklis aktyvina promotoriaus panaudojimą, kontroliuodamas transkripcijos iš to konkretaus promotoriaus efektyvumą ir greitį. Stiprintojai suaktyvina tik gulinčius-promotorių padėtis (t. y. promotoriai toje pačioje chromosomoje), tačiau jie gali veikti dideliais atstumais. Be to, jie gali būti ne tik 5 colių geno pusėje, bet ir kitoje DNR grandinėje (Maniatis ir kt., 1987). Genų, kurie transkribuoja santykinai didelius mRNR kiekius, promotoriai turi panašią struktūrą. Juose yra seka AΤA (kartais vadinama TATA dėžute arba, esantis dar toliau 5" pusėje. Prieš srovę esantis promotoriaus elementas dažniausiai yra CAAT sekos variantas, tačiau buvo nustatyti ir kiti promotoriaus elementai (Grosschedel ir Birnstiel, 1980; McKnight ir Tjian, 1986) (12.6 pav.).
Pirmą kartą β-globino geno promotorius buvo tiriamas atliekant eksperimentus, siekiant patikrinti specifinę klonuotos DNR transkripciją. Klonuoti genai gali būti teisingai transkribuoti, kai jie įvedami į varlių oocitų ar fibroblastų branduolius arba kai jie yra inkubuojami su išgryninta RNR polimeraze, esant supernatantiniams nukleotidams (Wasylyk ir kt., 1980). Patvirtinus geno transkripciją, restrikcijos fermentai naudojami specifiniams to geno ar aplinkinių regionų padalijimams gaminti. Tada galima nustatyti, ar modifikuotas genas ir toliau tinkamai transkribuojamas. Šių tyrimų rezultatai parodė, kad pirmųjų 109 bazių porų, esančių prieš dangtelio vietą, pakanka maksimaliai ß-globino geno transkripcijai (Grosveld ir kt., 1982; Dierks ir kt., 1983).
Kiti tyrėjai paaiškino šią išvadą klonuodami pelės globino geno sritį nuo 106-osios bazių poros prieš srovę (nuo 5 colių pusės) nuo transkripcijos pradžios (-106 padėtis) iki 475-osios bazinės poros (+475 padėtis) pirmasis egzonas (Myers ir kt., 1986), šie klonai buvo mutagenizuoti in vitro globino geno promotoriaus srityje.
Transkripcija yra RNR sintezė DNR šablone. Prokariotuose visų trijų tipų RNR sintezę katalizuoja vienas kompleksinis baltymų kompleksas – RNR polimerazė.
MRNR sintezė prasideda nuo RNR polimerazės atradimo specialioje DNR molekulėje, kuri nurodo vietą, kur prasideda transkripcija. propaguotojas Prisijungusi prie promotoriaus, RNR polimerazė išvynioja gretimą DNR spiralės posūkį. Šiuo metu dvi DNR grandinės išsiskiria, o vienoje iš jų fermentas sintezuoja mRNR. Ribonukleotidų surinkimas į grandinę vyksta atsižvelgiant į jų komplementarumą DNR nukleotidams, taip pat antiparaleliškai DNR šablono grandinės atžvilgiu. RNR polimerazė gali surinkti polinukleotidą tik nuo 5 colių galo iki 3 colio galo, tik viena iš dviejų DNR grandinių gali būti transkripcijos šablonas, būtent ta, kuri yra nukreipta į fermentą su 3 colių galu (3 colių →); 5") Ši grandinė vadinama kodogenine.
Terminatorius- tai sritis, kurioje sustoja tolesnis RNR grandinės augimas ir ji išsiskiria iš DNR šablono. RNR polimerazė taip pat atsiskiria nuo DNR, kuri atkuria jos dvigrandę struktūrą.
DNR molekulės fragmentas, įskaitant promotorių, transkribuotą seką ir terminatorių, sudaro transkripcijos vienetą - transkripcija.
Operono reguliavimas (t. y. reguliavimas transkripcijos lygiu) yra pagrindinis mechanizmas, reguliuojantis genų aktyvumą prokariotuose ir bakteriofaguose.
Operonas - genetinės medžiagos sekcija, kurios transkripcija vykdoma vienai RNR molekulei, kontroliuojant represoriaus baltymą.
Operonas susideda iš glaudžiai susijusių struktūrinių genų, koduojančių baltymus (fermentus), kurie vykdo nuoseklius metabolito biosintezės etapus. Kiekviename operone yra: promotorius, operatorius ir terminatorius.
Operatorius- nukleotidų seka, kuri jungiasi represoriaus baltymas ir neigiamai reguliuoja transkripcija kaimyninis genas. Operatorius yra tarp promotoriaus ir struktūrinių genų. Jis gali būti susijęs su specialiu baltymu – represoriumi, kuris neleidžia RNR polimerazei judėti DNR grandine ir trukdo fermentų sintezei. Taigi, genai gali būti įjungti ir išjungti, priklausomai nuo atitinkamų represorių baltymų buvimo ląstelėje.
Represorius- reguliacinis baltymas, kuris slopina jo reguliuojamo operono genų transkripciją dėl prisijungimo prie operatoriaus (operono reguliavimo vieta). Dėl to nutrūksta atitinkamos mRNR ir, atitinkamai, operono koduojamų fermentų sintezė. Represorius yra sintetinamas kontroliuojant genų reguliatorių nuo 10 iki 20 molekulių vienoje ląstelėje aktyvių, t.y. galinčių tiesiogiai prisijungti prie operatorių, arba neaktyvių formų. Aktyvaus represoriaus susidarymas būdingas indukuojamiems fermentams, kurių sintezė prasideda tik tada, kai į ląstelę patenka specifinės mažos molekulinės masės medžiagos – induktoriai. . Induktorius- maža efektorinė molekulė, kuri jungiasi su reguliuojančiu baltymu, arba fizinis veiksnys (šviesa, temperatūra), skatinantis neaktyvios būsenos genų ekspresiją.
Geno 5' srities promotoriaus-reguliavimo seka, kuri lemia RNR polimerazės prisijungimo prie DNR vietą. Promotoriuje yra dvi sekos, kurios vaidina svarbų vaidmenį inicijuojant: TATA dėžė ir CCAAT domenas. Promotorius nurodo RNR polimerazei, kur pradėti sintezę.
102 klausimas. Paaiškinkite sąvokas: nuorašas ir nuorašas.
Nuorašas– RNR molekulė, susidaranti dėl DNR dalies transkripcijos
Transkripcija- visų vienos ląstelės sintezuotų transkriptų visuma, įskaitant mRNR ir nekoduojančią RNR
Klausimas: Kokias molekules sintetina RNR polimerazė I?
Sintetina 5.8srRNR, 18srRNR, 28srRNR
atsakingas už didelių rRNR sintezę, jis yra lokalizuotas branduolyje.
Klausimas: Kokias molekules sintetina RNR polimerazė II?
Sintetina mRNR, snRNR (mažo branduolio), miRNR, miRNR (mažai trukdančią),
mRNR, ji yra lokalizuota citoplazmoje
Klausimas: Kokias molekules sintetina RNR polimerazė III.
Sintetina 5sRNR, tRNR, snRNR dalį, mažąją rRNR
Klausimas: Ką reiškia transkripcijos mechanizmas (RNR sintezė DNR šablone).
Dėl transkripcijos susidaro pirminė transkriptoRNR.
107 klausimas: kas vadinamas RNR apdorojimu? Įvardykite, iš kokių atskirų procesų jis susideda.
MRNR brendimo procesas vadinamas apdorojimu. Uždengimas, poliadenilinimas, sujungimas
Klausimas: kokia yra RNR transkriptų sujungimo reakcija? Splaisingosomos vaidmuo šiame procese.
Sujungimas- Tai intronų išpjovimas iš mRNR molekulės ir egzonų susiuvimas naudojant ligazės fermentus.
Nustatytas makromolekulinis procesas apima daug makromolekulių. Atliekama stambiamolekulinė struktūra (splicingosoma->mažoji branduolinė RNR). snRNR atpažįsta ribą tarp egzonų ir intronų ir prie jų prisijungia.
109 klausimas. Kada įvyksta eukariotų RNR 5′ galo uždengimas ir ką tai reiškia? Nuorašai, kurių RNR polimerazės yra apribotos.
Iš karto po transkripcijos. Metilinta guanozino liekana yra prijungta prie 5" mRNR galo; ši struktūra vadinama dangteliu. Dangtelis palengvina mRNR prisijungimą prie ribosomos citoplazmoje.
Tik RNR polimerazės II transkriptai
Klausimas: Išvardykite visas mRNR ribojimo reikšmės nuostatas.
Užtikrina tolesnės transkripcijos efektyvumą.
Apsaugo nuorašą nuo 5'-egzonukleazių (5'-5' jungčių) degradacijos
Būdas tolesniam procesui: skatina 3'-poliadenilinimą ir sujungimą.
Reikalingas mRNR eksportui iš branduolio
Užtikrina mRNR prisijungimą prie ribosomos citoplazmoje
111.Kas yra mRNR 3′ galo poliadenilinimas. Kokį vaidmenį atlieka šis procesas? Nuorašai, kurių RNR polimerazės patiria šį procesą.
Poliadenilinimas– į 3" mRNR molekulės galą pridedama nuo 100 iki 200 adenilo nukleotidų. Susidaro poli-A sritis. Ši sritis stabilizuoja mRNR molekulę ir skatina jos išsiskyrimą iš branduolio į citoplazmą.
RNR nuorašai- polimerazė II
Kas yra alternatyvus sujungimas? Kokią biologinę reikšmę turi ši sujungimo forma?
alternatyvus sujungimas yra sujungimo forma, kai iRNR brendimo metu egzonų susijungimas vyksta įvairiais deriniais. Tokiu atveju egzonų tvarka nepažeidžiama. Sujungimas užtikrina, kad genai koduotų skirtingus baltymus, o tai yra eukariotų baltymų įvairovės mechanizmas (daugiau nei 70 % žmogaus genų atlieka alternatyvų sujungimą).