ARN -polimeraza este o enzimă care sintetizează moleculele de ARN. Într-un sens restrâns, ARN polimeraza este de obicei numită ARN polimeraze dependente de ADN care sintetizează molecule de ARN pe un șablon de ADN, adică realizează transcripția. Enzimele din clasa ARN polimerazei sunt foarte importante pentru funcționarea celulei, așa că se găsesc în toate organismele și în multe virusuri. Din punct de vedere chimic, ARN polimerazele sunt nucleotidil transferaze care polimerizează ribonucleotidele la capătul de 3" al lanțului de ARN.
Un promotor este o secvență de nucleotide ADN recunoscută de ARN polimerază ca punct de plecare pentru transcrierea specifică sau semnificativă. La procariote, promotorul include un număr de motive care sunt importante pentru recunoașterea sa de către ARN polimerază, în special așa-numitele secvențe -10 și -35. Promotorul este asimetric, ceea ce permite ARN polimerazei să înceapă transcripția în direcția corectă și indică care dintre cele două catene de ADN va servi ca șablon pentru sinteza ARN.
Regiunea promotor din operon se poate suprapune parțial sau deloc cu regiunea operator a cistronului (genei).
Promotorul sub care se află regiunea ADN-ului care codifică ARN joacă un rol decisiv în intensitatea expresiei acestei gene în fiecare tip de celulă specific. Activarea promotorului este determinată de prezența unui set diferit de factori de transcripție în fiecare tip de celulă.
Terminator este o secvență de nucleotide ADN recunoscută de ARN polimerază ca un semnal pentru oprirea sintezei moleculei de ARN și disocierea complexului de transcripție.
Adesea, aceste secvențe se termină cu un lanț de nucleotide de timină (în transcrieri - uridină), care este precedat de o regiune care conține secvențe interne, reciproc complementare, în orientări opuse (structuri „ac de păr”). Regiunea ac de păr este îmbogățită cu perechi GC, oferind acestei structuri o stabilitate mai mare.
78. Diferențele în procesul de transcripție în diferite celule. Structura mozaică a genelor. Sinteza ARN heteronuclear, conversia lui în ARN mesager. Îmbinare. Prelucrare.
Unii virusuri (cum ar fi HIV, care cauzează SIDA) au capacitatea de a transcrie ARN-ul în ADN. HIV are un genom ARN care este integrat în ADN. Ca rezultat, ADN-ul virusului poate fi combinat cu genomul celulei gazdă. Principala enzimă responsabilă de sintetizarea ADN-ului din ARN se numește reversease. Una dintre funcțiile revertasezei este de a crea ADN complementar (ADNc) din genomul viral. Enzima asociată ribonucleaza H scindează ARN-ul, iar reversease sintetizează ADNc din dubla helix ADN. ADNc este integrat în genomul celulei gazdă prin integrază. Rezultatul este sinteza proteinelor virale de către celula gazdă, care formează noi viruși. În cazul HIV, este programată și apoptoza (moartea celulară) a limfocitelor T. În alte cazuri, celula poate rămâne un răspânditor de viruși.
Unele celule eucariote conțin enzima telomeraza, care prezintă și activitate de transcripție inversă. Cu ajutorul lui, se sintetizează secvențe repetate în ADN. Telomeraza este adesea activată în celulele canceroase pentru a duplica la nesfârșit genomul fără a pierde secvența de ADN care codifică proteine.
O trăsătură distinctivă a structurii multor gene eucariote este structura mozaică (discontinuă) a părții semantice a genelor.
Această diferență este asociată cu alternanța regiunilor semantice care transportă informații despre secvența de aminoacizi din proteină - exoni și secțiuni de secvențe necodante care sunt semnificativ mai lungi decât exonii - introni. Ca rezultat, lungimea totală a genei se dovedește a fi de aproximativ 5-7 ori mai mare decât ne-am aștepta.
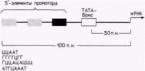
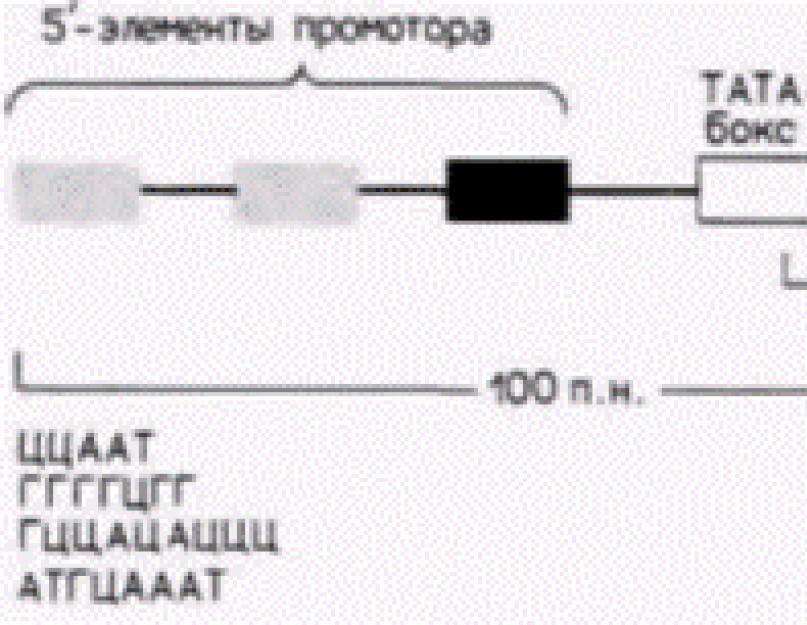
La începutul genei (înaintea părții sale semantice) există zone care asigură o reglare adecvată a genei. Ele sunt împărțite în două grupe: nespecifice (același pentru toate genele) și specifice (caracteristice doar pentru o anumită genă). Regiunile de reglare nespecifice la eucariote sunt numite „TATA-BOX”. Ele alternează între timină și adenină. Această regiune se află la 30 de nucleotide la stânga începerii citirii genei. S-a stabilit că ARN polimeraza se sprijină pe ADN în așa fel încât partea sa de recunoaștere închide TATA BOX, iar centrul activ al ARN polimerazei apare deasupra primei nucleotide care trebuie citită. Aceasta este urmată de regiunea promotor, constând dintr-un situs de recunoaștere, un situs de legare și un situs de inițiere a transcripției (A sau D). Combinația de nucleotide din promotor este astfel încât, dacă cadrul de citire este setat incorect, se obține o secvență caracteristică codonilor de oprire și transcripția se oprește. Regiunea promotoare este urmată de un palindrom („flip”) sau repetare inversată. Aceasta este o secvență de ADN dublu catenar care citește la fel în ambele direcții („Cazacul” - de la dreapta la stânga și de la stânga la dreapta citește la fel). Uneori, o astfel de secvență este numită „regiune dublu simetrică”, unde axa de simetrie este punctul central în raport cu care secvența rămâne aceeași. O proprietate importantă a palindromilor este capacitatea de a forma structuri în ac de păr în ARN sau structuri încrucișate în ADN. Dacă cadrul de citire este instalat incorect, palindromul ADN se transformă într-o „cruce”, ceea ce face imposibilă progresul ulterioar al enzimei. Uneori, palindromii pot fi localizați la sfârșitul unei gene după partea sa semantică și, de asemenea, formând o „cruce”, vor opri sinteza ARNm. Partea semantică a genei, așa cum am scris mai sus, constă în secvențe alternante intron-exon. Partea structurală a genei conține un promotor și un palindrom.
Recent, au fost identificate incluziuni de reglare a genelor specifice - amplificatori. Ele sunt situate în fața genei la o distanță de sute și mii de perechi de nucleotide. Eucariotele au proteine reglatoare speciale care recunosc intensificatorul și se atașează de acesta. În acest fel, gena este activată. Genomul este format nu numai din gene nucleare, ci și din gene citoplasmatice. În genom, genele îndeplinesc diferite funcții. În funcție de funcțiile pe care le îndeplinesc, genele sunt împărțite în gene obligatorii ele codifică funcții vitale în celule de toate tipurile (uneori numite gene „de menaj”, gene structurale, gene reglatoare și gene de punctuație; În timp ce ARNm-urile procariote (bacteriene și arheale), cu rare excepții, sunt imediat gata pentru traducere și nu necesită procesare specială, pre-ARNm-urile eucariote suferă modificări intensive. Astfel, simultan cu transcripția, are loc „editarea” (splicing) pe porțiunea deja sintetizată a ARNm. În timpul procesului de splicing, secvențele care nu codifică proteine - introni - sunt îndepărtate din pre-ARNm, se adaugă o nucleotidă specială modificată (capac) la capătul de 5" al moleculei și mai multe adenine, așa-numita coadă de poliadenină, sunt adăugate la capătul de 3". Capacul este recunoscut de factorii de inițiere, proteinele responsabile de atașarea la ARNm-ul ribozomului, iar coada poliadeninei se leagă de o proteină specială, PAB. De obicei, aceste modificări post-transcripționale ale ARNm eucariotic sunt denumite „procesare ARNm”. Poliadenilarea este necesară pentru transportul majorității ARNm în citoplasmă și protejează moleculele de ARNm de degradarea rapidă (mărește timpul de înjumătățire al acestora). Moleculele de ARNm lipsite de o regiune poli-A (de exemplu, cele virale) sunt distruse rapid în citoplasma celulelor eucariote de către ribonucleaze.
Splicing (din limba engleză splice - a splice sau a lipi capetele ceva) este procesul de tăiere a anumitor secvențe de nucleotide din moleculele de ARN și de unire a secvențelor care rămân în molecula „matură” în timpul procesării ARN. Acest proces are loc cel mai adesea în timpul maturării ARN-ului mesager (ARNm) la eucariote, timp în care, prin reacții biochimice care implică ARN și proteine, se elimină secțiuni ale ARNm care nu codifică o proteină (introni) și secțiuni care codifică amino. secvența acidă - exonii sunt legați între ei. Astfel, pre-ARNm imatur este convertit în ARNm matur, din care sunt citite (traduse) proteinele celulare. Cele mai multe gene care codifică proteine procariote nu au introni, astfel încât splicingul pre-ARNm este rară în ele. Îmbinarea ARN-urilor de transfer (ARNt) și a altor ARN-uri necodificatoare apare și la reprezentanții eucariotelor, bacteriilor și arheilor.
procesarea ARN
Între transcripție și traducere, molecula de ARNm suferă o serie de modificări secvențiale care asigură maturarea matricei funcționale pentru sinteza lanțului polipeptidic. Un capac este atașat la capătul 5΄, iar o coadă poli-A este atașată la capătul 3΄, ceea ce crește durata de viață a ARNm. Odată cu apariția procesării în celula eucariotă, a devenit posibilă combinarea exonilor genici pentru a obține o varietate mai mare de proteine codificate de o singură secvență de nucleotide ADN - splicing alternativ.
Pentru a realiza transcrierea corectă, sunt necesare două tipuri de elemente de reglementare. Elementele de reglementare de primul tip sunt numite cis-regulatori. Sunt secvențe specifice de ADN pe un anumit cromozom. Cis -regulatorii actioneaza numai asupra genelor din apropiere. Al doilea tip este numit trans-regulatori. Acestea sunt molecule solubile (inclusiv proteine și ARN) care sunt produse de o genă și interacționează cu alte gene de pe același cromozom sau de pe alți cromozomi. Dacă ne întoarcem la inducerea genelor în lac -operonE., coli Acestea sunt molecule solubile (inclusiv proteine și ARN) care sunt produse de o genă și interacționează cu alte gene de pe același cromozom sau de pe alți cromozomi. Dacă ne întoarcem la inducerea genelor în apoi ne putem aminti că gena represoare produce o proteină represoare care interacționează cu secvența operatorului pentru gene -operon.În acest caz operatorul este Acestea sunt molecule solubile (inclusiv proteine și ARN) care sunt produse de o genă și interacționează cu alte gene de pe același cromozom sau de pe alți cromozomi. Dacă ne întoarcem la inducerea genelor în- cis -un element de reglementare, deoarece doar controlează operonul propriului cromozom. -operon.(O secvență de operator mutant pe alt cromozom poate atașa sau nu o proteină represoare.) În schimb, o proteină represoare este
transă -operon.-regulator. deoarece este produs de un cromozom și se leagă de -operator reglator pe alt cromozom (Fig. 12.5). situat de obicei imediat înainte de locul unde începe
Gilbert s. Biologia dezvoltării: în 3 volume T. 2: Trad. Din engleză – M.: Mir, 1994. – 235 p.
142 _______________ CAPITOLUL 12 _____________________________________________________________________________
|
Orez. 12.5. Schema de reglare diferențială a genelor în E. E.; arătat -operon.- Și -un element de reglementare, deoarece doar controlează-elementele de reglementare. În celulele de tip sălbatic, starea inductibilă este caracterizată prin faptul că ARN-ul pentru β-galactozidază nu este transcris până când lactoza este prezentă. În absența lactozei, proteina represoare (R) este codificată de genă i, se alătură site-ului web al operatorului ( O), inhibând astfel transcripția de către ARN polimeraza de la promotor ( p). E. Dacă este prezentă lactoza, aceasta se leagă de proteina represoare, ca urmare represorul nu se poate lega de ADN și transcripția continuă. Natura solubilă a acestui represor a fost demonstrată în experimente pe mutanți. Coli i – Când celulele bacteriene haploide care poartă gena i, devin parțial diploid cu gena i + tip sălbatic ( -un element de reglementare, deoarece doar controlează), este sintetizat un represor de tip sălbatic, care este capabil să facă inductibilă gena ß-galactozidazei originale. -operon. Această proteină represoare este |
|
|
|
-element de reglementare. Secvențele promotorului și operatorului sunt |

-elementele de reglementare. -operon. Orez. 12.6. Un promotor tipic pentru o genă care codifică proteine eucariote. Gena prezentată conține o cutie TATA și trei elemente promotoare de 5". Exemple de astfel de elemente 5' sunt prezentate în partea de jos a figurii. (După Maniatis și colab., 1987.)
transcripție și au aproximativ 100 de perechi de baze în lungime. Regiunea promotor este necesară pentru legarea ARN polimerazei II și inițierea precisă a transcripției. Amplificatorul activează utilizarea promotorului prin controlul eficienței și ratei transcripției de la acel promotor particular. Potenziatorii activează numai pe cei care zac înăuntru-pozitia promotorilor (adica promotori pe acelasi cromozom), dar pot functiona pe distante mari. În plus, ele pot fi localizate nu numai pe partea de 5" a genei, ci și pe o altă catenă de ADN (Maniatis și colab., 1987). Promotorii genelor care transcriu cantități relativ mari de ARNm au structuri similare. Ele conțin secvența AΤA (uneori numită cutie TATA sau, situat chiar mai departe pe partea de 5". Elementul promotor din amonte este de obicei o variație a secvenței CAAT, dar au fost identificate și alte elemente promotoare (Grosschedl și Birnstiel, 1980; McKnight și Tjian, 1986) (Fig. 12.6).
Pentru prima dată, promotorul genei β-globinei a fost studiat în experimente pentru a testa transcripția specifică a ADN-ului clonat. Genele clonate pot fi transcrise corect atunci când sunt introduse în nucleele ovocitelor de broaște sau fibroblastelor sau când sunt incubate cu ARN polimerază purificată în prezența nucleotidelor supernatante (Wasylyk și colab., 1980). Odată ce transcripția unei gene a fost confirmată, enzimele de restricție sunt utilizate pentru a produce diviziuni specifice în acea genă sau în regiunile învecinate. Apoi se poate determina dacă gena modificată continuă să fie transcrisă corect. Rezultatele acestor studii au arătat că primele 109 perechi de baze care preced situsul cap sunt suficiente pentru transcripția maximă a genei ß-globinei (Grosveld și colab., 1982; Dierks și colab., 1983).
Alți cercetători au clarificat această concluzie prin clonarea unei regiuni a genei globinei de șoarece din a 106-a pereche de baze în amonte (din partea de 5") a începutului transcripției (poziția -106) până la a 475-a pereche de baze (poziția +475) în primul exon (Myers et al., 1986). Aceste clone au fost mutagenizate in vitro în regiunea promotoare a genei globinei.
Transcriere este sinteza ARN-ului pe un model de ADN. La procariote, sinteza tuturor celor trei tipuri de ARN este catalizată de un complex proteic complex - ARN polimeraza.
Sinteza ARNm începe cu descoperirea de către ARN polimerază a unei regiuni speciale din molecula de ADN, care indică locul unde începe transcripția - promotor După legarea de promotor, ARN polimeraza desfășoară tura adiacentă a helixului ADN. Două catene de ADN diverg în acest punct, iar pe una dintre ele enzima sintetizează ARNm. Asamblarea ribonucleotidelor într-un lanț are loc în conformitate cu complementaritatea lor cu nucleotidele ADN și, de asemenea, antiparalel în ceea ce privește catena matriță ADN. ARN polimeraza este capabilă să asambleze o polinucleotidă doar de la capătul de 5" la capătul de 3" doar una dintre cele două catene de ADN poate servi ca matriță pentru transcripție, și anume cea care se află în fața enzimei cu capătul său de 3" (3" →; 5") Acest lanț se numește codogenic.
Terminator- aceasta este zona în care creșterea ulterioară a lanțului de ARN se oprește și este eliberată din matrița de ADN. ARN polimeraza se separă și de ADN, care îi restabilește structura dublu catenară.
Un fragment al unei molecule de ADN, incluzând un promotor, o secvență transcrisă și un terminator, formează o unitate de transcripție - transcriere.
Reglarea operonilor (adică reglarea la nivel transcripțional) este principalul mecanism de reglare a activității genelor la procariote și bacteriofagi.
Operon - o secțiune de material genetic, a cărei transcriere se realizează pe moleculă de ARN sub controlul unei proteine represoare.
Un operon constă din gene structurale strâns legate care codifică proteine (enzime) care realizează etape succesive ale biosintezei unui metabolit. Fiecare operon conține: un promotor, un operator și un terminator.
Operator- secvența de nucleotide care se leagă proteina represoareși reglarea negativă transcriere vecine gena. Operatorul este situat între promotor și genele structurale. Poate fi asociat cu o proteină specială - un represor, care împiedică ARN polimeraza să se deplaseze de-a lungul lanțului ADN și împiedică sinteza enzimelor. Astfel, genele pot fi activate și oprite în funcție de prezența proteinelor represoare corespunzătoare în celulă.
Represor- o proteină reglatoare care suprimă transcrierea genelor operonului pe care îl reglează ca urmare a legării de operator (situsul de reglare al operonului). Aceasta duce la încetarea sintezei ARNm-ului corespunzător și, în consecință, a enzimelor codificate de operon. Represorul este sintetizat sub controlul regulatorului genei în cantitate de 10 până la 20 de molecule per celulă sub formă de forme active, adică capabile să se lege direct de operator, sau forme inactive. Formarea unui represor activ este caracteristică enzimelor inductibile, a căror sinteză începe numai atunci când anumite substanțe cu molecul scăzut - inductori - intră în celulă. . Inductor- o moleculă efectoră mică care se leagă de o proteină reglatoare sau de un factor fizic (lumină, temperatură) care stimulează expresia genelor aflate în stare inactivă.
Secvența de reglare promotor a regiunii 5’ a genei, care determină locul de atașare a ARN polimerazei la ADN. Promotorul conține două secvențe care joacă un rol important în inițiere: caseta TATA și domeniul CCAAT. Promotorul spune ARN polimerazei de unde să înceapă sinteza.
102 intrebare. Explicați conceptele: transcriere și transcriptom.
Transcriere– o moleculă de ARN formată ca urmare a transcripției unei secțiuni de ADN
Transcriere-totalitatea tuturor transcrierilor sintetizate de o celulă, include ARNm și ARN necodant
Întrebare: Ce molecule sunt sintetizate de ARN polimeraza I?
Sintetizează 5,8srARN, 18srARN, 28srARN
responsabil de sinteza ARNr-urilor mari, este localizat în nucleol.
Întrebare: Ce molecule sunt sintetizate de ARN polimeraza II?
Sintetizează ARNm, snRNA (nuclear scăzut), miARN, miARN (cu interferență scăzută),
ARNm, este localizat în citoplasmă
Întrebare: Ce molecule sunt sintetizate de ARN polimeraza III.
Sintetizează 5sARN, ARNt, o parte din ARNsn, ARNr mic
Întrebare: La ce se rezumă mecanismul de transcripție (sinteza ARN pe un șablon de ADN).
Ca rezultat al transcripției, se formează un transcriptiARN primar.
Întrebarea 107: Ce se numește procesarea ARN? Numiți din ce procese individuale constă.
Procesul de maturare a ARNm se numește procesare. Capacul, poliadenilarea, îmbinare
Întrebare: Care este reacția de splicing a transcriptelor ARN? Rolul splingozomului în acest proces.
Îmbinare- Aceasta este tăierea intronilor dintr-o moleculă de ARNm și cusătura exonilor folosind enzime ligază.
Procesul macromolecular stabilit implică un număr mare de macromolecule. Se realizează structura macromoleculară (splicingosome->ARN nuclear mic). ARNsn recunosc granița dintre exoni și introni și se leagă de ei.
Întrebarea 109: Când are loc limitarea capătului 5′ al ARN-ului la eucariote și ce înseamnă? Transcrieri ale căror ARN polimeraze sunt supuse plafonării.
Imediat după transcriere. Un reziduu de guanozină metilat este atașat la capătul de 5" al ARNm; această structură se numește capac. Capacul facilitează legarea ARNm la ribozomul din citoplasmă.
Doar transcriptele ARN polimerază II
Întrebare: Enumerați toate prevederile privind semnificația plafonării ARNm.
Asigură eficiența transcripției ulterioare.
Protejează transcriptul de degradarea de către 5’-exonucleaze (legături 5’-5’)
O modalitate de a continua procesul: stimulează 3’-poliadenilarea și îmbinarea.
Necesar pentru exportul de ARNm din nucleu
Oferă legarea ARNm la ribozomul din citoplasmă
111.Ce este poliadenilarea capătului 3′ al ARNm. Ce rol are acest proces? Transcrieri ale căror ARN polimeraze suferă acest proces.
Poliadenilare– se adaugă de la 100 la 200 de nucleotide adenil la capătul de 3" al moleculei de ARNm. Se formează o regiune poli-A. Această regiune stabilizează molecula de ARNm și promovează eliberarea acesteia din nucleu în citoplasmă.
transcriptele ARN-polimeraza II
Ce este splicing alternativ? Ce semnificație biologică are această formă de îmbinare?
splicing-ul alternativ este o formă de splicing în care unirea exonilor în timpul maturării ARNm are loc în diferite combinații. În acest caz, ordinea exonilor nu este încălcată. Splicing-ul asigură că genele codifică diferite proteine, care este un mecanism de diversitate a proteinelor la eucariote (mai mult de 70% dintre genele umane suferă splicing alternativ).