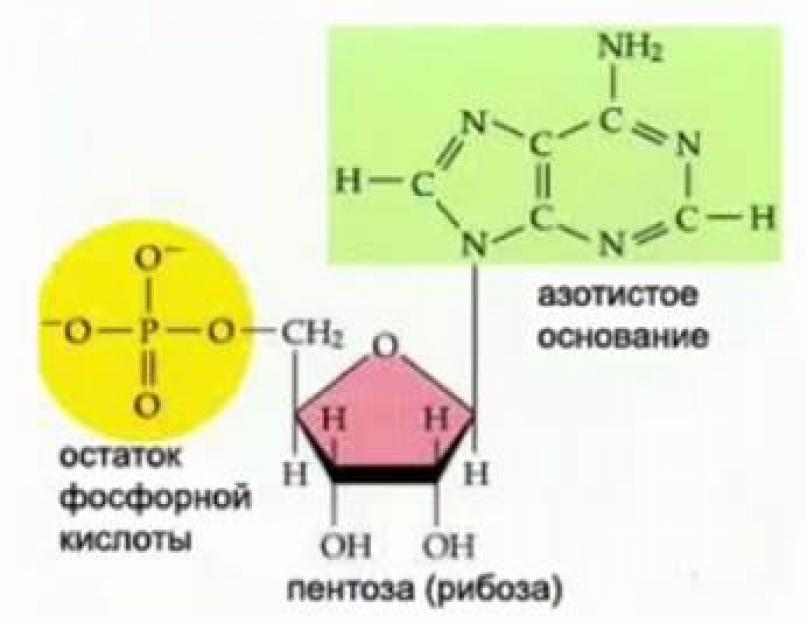
Nükleotidler içindeki RNA monomerleri beş karbonlu bir şeker (pentoz), fosforik asit (bir fosforik asit kalıntısı) ve bir nitrojenli baz içerir (bkz. Şekil 2).

Pirinç. 2. RNA nükleotidinin yapısı
RNA'nın azotlu bazları urasil, sitozin, adenin ve guanindir. RNA nükleotid monosakarit ribozdur (bkz. Şekil 2).
RNA tek sarmallı bir moleküldür daha küçük boyutlar bir DNA molekülünden daha fazlasıdır.
Bir RNA molekülü 75 ila 10.000 nükleotid içerir.

Pirinç. 3. RNA virüsü
İnfluenza virüsü gibi birçok virüs, tek nükleik asitleri olarak bir RNA molekülü içerir (bkz. Şekil 3). İnsanlar için patojenik olan, DNA içerenlerden daha fazla RNA içeren virüs vardır. Çocuk felcine, hepatit A'ya ve akut soğuk algınlığına neden olurlar.
Arbovirüsler eklembacaklılar tarafından taşınan virüslerdir. Bunlar kene kaynaklı ve Japon ensefalitinin yanı sıra sarı hummanın da etken maddeleridir.
İnsan solunum ve bağırsak hastalıklarının nadir patojenleri olan Reovirüsler (bkz. Şekil 4), genetik materyallerinin çift sarmallı bir RNA molekülü formunda sunulması nedeniyle özel bilimsel ilgi konusu haline gelmiştir.

Pirinç. 4. Reovirüsün yapısı
Ayrıca birçok hastalığa neden olan retrovirüsler de vardır. onkolojik hastalıklar.
Gerçekleştirilen yapıya ve fonksiyona bağlı olarak üç ana RNA türü ayırt edilir: ribozomal, taşıma ve bilgilendirici (şablon).
1. Haberci RNA'sı
Araştırmalar haberci RNA'nın hücredeki toplam RNA içeriğinin %3-5'ini oluşturduğunu göstermiştir. Bu, DNA molekülünün zincirlerinden birinde transkripsiyon sırasında oluşan tek iplikli bir moleküldür. Bunun nedeni, nükleer organizmalarda DNA'nın çekirdekte yer alması ve protein sentezinin sitoplazmadaki ribozomlarda gerçekleşmesi nedeniyle bir "aracıya" ihtiyaç duyulmasıdır. Bir "aracının" işlevi, haberci RNA tarafından gerçekleştirilir; proteinin yapısı hakkındaki bilgileri, DNA'nın bulunduğu hücre çekirdeğinden, bu bilginin uygulandığı ribozomlara iletir (bkz. Şekil 5).

Pirinç. 5. Haberci RNA (mRNA)
Kopyalanan bilgi miktarına bağlı olarak haberci RNA molekülü farklı uzunluklara sahip olabilir.
Haberci RNA'ların çoğu hücrede kısa bir süre için bulunur. Bakteri hücrelerinde bu tür RNA'ların varlığı dakikalarla belirlenirken, memeli hücrelerinde (kırmızı kan hücrelerinde) hemoglobin (protein) sentezi, kırmızı kan hücrelerinin çekirdeklerini kaybetmesinden sonra birkaç gün daha devam eder.
2. Ribozomal RNA
Ribozomal RNA'lar (bkz. Şekil 6), hücrede bulunan tüm ribozomların %80'ini oluşturur. Bu RNA'lar nükleolusta sentezlenir ve hücrede sitoplazmada bulunurlar, burada proteinlerle birlikte ribozomlar oluştururlar. Protein sentezi ribozomlarda gerçekleşir. Burada haberci RNA'nın içerdiği "kod", bir protein molekülünün amino asit dizisine çevrilir.

Pirinç. 6. Ribozomal RNA (rRNA)
3. RNA'yı aktar
Transfer RNA'ları (bkz. Şekil 7), DNA üzerindeki çekirdekte oluşturulur ve daha sonra sitoplazmaya doğru hareket eder.

Pirinç. 7. Transfer RNA'sı (tRNA)
Bu RNA'lar hücredeki toplam RNA içeriğinin yaklaşık %10'unu oluşturur. 80-100 nükleotidlik en kısa moleküllere sahiptirler.
Transfer RNA'ları bir amino asit bağlar ve onu protein sentezi bölgesine, yani ribozomlara taşır.
Bilinen tüm transfer RNA'lar, nitrojenli bazlar arasındaki tamamlayıcı etkileşimler nedeniyle, şekil olarak yonca yaprağına benzeyen ikincil bir yapı oluşturur (bkz. Şekil 8). Bir tRNA molekülünün iki aktif bölgesi vardır; bir ucunda üçlü bir antikodon ve diğer ucunda bir amino asidi bağlayan bir alıcı bölge.

Pirinç. 8. tRNA’nın yapısı (“yonca yaprağı”)
Her amino asit, üç nükleotidin karşılık gelen bir kombinasyonuna sahiptir. üçlü.

Pirinç. 9. Genetik kod tablosu
Amino asit kodlama üçlüleri - kodonlar DNA (bkz. Şekil 9) - mRNA'nın üçlüleri (kodonları) bilgisi şeklinde aktarılır. tRNA yonca yaprağının tepesinde, karşılık gelen mRNA kodonuna tamamlayıcı olan bir üçlü nükleotid bulunur (bkz. Şekil 10). Bu üçlü, farklı amino asitler taşıyan tRNA'lar için farklıdır ve tam olarak bu tRNA'nın taşıdığı amino asidi kodlar. İsmini aldı antikodon.

Pirinç. 10.tRNA
Alıcı uç, belirli bir amino asit için "iniş alanıdır".
Böylece, çeşitli türler RNA'lar, uygulamayı amaçlayan tek bir fonksiyonel sistemi temsil eder. kalıtsal bilgi protein sentezi yoluyla.
RNA dünyası kavramı, çok uzun zaman önce, bir RNA molekülünün hem DNA molekülünün hem de proteinlerin işlevini yerine getirebildiğine dayanmaktadır.
Canlı organizmalarda hemen hemen tüm işlemler protein enzimleri sayesinde gerçekleşir. Ancak proteinler kendi kendilerini kopyalayamazlar ve DNA'nın içerdiği bilgilere dayanarak hücrelerde sentezlenirler. Ancak DNA'nın iki katına çıkması yalnızca proteinlerin ve RNA'nın katılımı nedeniyle gerçekleşir. Sonuç olarak, yaşamın kökeni teorisi çerçevesinde böyle bir şeyin kendiliğinden ortaya çıkması nedeniyle bir kısır döngü oluşur. karmaşık sistem pek olası değil.
1980'lerin başında, Check ve Altman bilim adamlarının laboratuvarında (sahipler Nobel Ödülü kimyada) RNA'nın katalitik yeteneği ABD'de keşfedildi. RNA katalizörleri adlandırıldı ribozimler(bkz. Şekil 11).

Pirinç. 11. Kataliz görevini yerine getiren RNA ribozim molekülünün yapısı
Ribozomların aktif merkezinin de içerdiği ortaya çıktı büyük sayı ribozomal RNA. RNA ayrıca çift sarmal oluşturma ve kendi kendini kopyalama yeteneğine de sahiptir. Yani, RNA tamamen özerk bir şekilde var olabilir, örneğin yeni ribonükleatidlerin sentezi gibi metabolik reaksiyonları katalize edebilir ve kendi kendini yeniden üreterek katalitik özelliklerini nesilden nesile koruyabilir. Rastgele mutasyonların birikmesi, daha etkili katalizörler olan belirli proteinlerin sentezini katalize eden RNA'ların ortaya çıkmasına neden oldu ve bu nedenle bu mutasyonlar sırasında sabitlendi. doğal seçilim. Genetik bilginin özel depoları da ortaya çıktı - DNA molekülü ve RNA, DNA ile proteinler arasında bir aracı haline geldi.
Referanslar
- Kamensky A.A., Kriksunov E.A., Pasechnik V.V. Genel biyoloji 10-11. sınıf Bustard, 2005.
- Biyoloji. 10. sınıf. Genel biyoloji. Temel seviye/ P.V. Izhevsky, O.A. Kornilova, T.E. Loschchilina ve diğerleri - 2. baskı, revize edildi. - Ventana-Graf, 2010. - 224 s.
- Belyaev D.K. Biyoloji 10-11 sınıf. Genel biyoloji. Temel seviye. - 11. baskı, stereotip. - M.: Eğitim, 2012. - 304 s.
- Agafonova I.B., Zakharova E.T., Sivoglazov V.I. Biyoloji 10-11 sınıf. Genel biyoloji. Temel seviye. - 6. baskı, ekleyin. - Bustard, 2010. - 384 s.
- Orgchem.ru ().
- Appteka.ru ().
- Youtube.com().
Ev ödevi
- 12. paragrafın sonundaki 4, 5. Sorular (s. 52) - Kamensky A.A., Kriksunov E.A., Pasechnik V.V. "Genel Biyoloji", 10-11. Sınıflar ()
- Nükleik asitler hücrede nerede bulunur?
Ribonükleik asit, DNA'da olduğu gibi fosfodiester köprüleri ile birbirine bağlanan purin ve pirimidin ribonükleotitlerin bir kopolimeridir (Şekil 37.6). Bu iki tip nükleik asitin pek çok ortak noktası olmasına rağmen, birçok yönden birbirlerinden farklıdırlar.
1. RNA'da purin veya pirimidin bazları ve fosfat gruplarının bağlandığı karbonhidrat kalıntısı 2-deoksiriboz değil (DNA'daki gibi) ribozdur.
2. RNA'nın pirimidin bileşenleri DNA'nınkinden farklıdır. RNA, DNA gibi, adenin, guanin ve sitozin nükleotidlerini içerir. Aynı zamanda RNA (aşağıda tartışacağımız bazı özel durumlar hariç) timin içermez; RNA molekülündeki yeri urasil tarafından işgal edilmiştir.
3. RNA tek sarmallı bir moleküldür (çift sarmallı bir yapıya sahip olan DNA'dan farklı olarak), ancak RNA zincirinde tamamlayıcı dizili (zıt polariteli) bölümler varsa, tek bir RNA zinciri katlanma yeteneğine sahiptir. çift sarmallı özelliklere sahip "saç tokası" adı verilen yapıları oluşturur ( Şekil 37.7).

Pirinç. 37.6. Pürin ve pirimidin bazlarının - adenin (A), urasil (U), sitozin (C) ve guanin (-) bir N- ile bağlanan ribozil kalıntılarını bağlayan bir fosfodiester omurgası tarafından tutulduğu bir ribonükleik asit (RNA) molekülünün bir fragmanı karşılık gelen nükleik bazlara glikosidik bağ Lütfen unutmayın: RNA zincirinin, 5- ve 3-terminal fosfat kalıntılarıyla gösterilen spesifik bir yönü vardır.
4. RNA molekülü, DNA iplikçiklerinden yalnızca birine tamamlayıcı tek iplikli olduğundan, içindeki guanin içeriğinin mutlaka sitozin içeriğine eşit olması gerekmez ve adenin içeriğinin mutlaka urasil içeriğine eşit olması gerekmez.
5. RNA, alkali ile mononükleotidlerin 2,3-siklik diesterlerine hidrolize edilebilir; hidrolizin ara ürünü, DNA'nın alkalin hidrolizi sırasında ikincisinde 2-hidroksil gruplarının bulunmaması nedeniyle oluşmayan 2,U,5-triesterdir; RNA'nın alkali kararsızlığı (DNA ile karşılaştırıldığında) kullanışlı özellik Hem teşhis hem de analitik amaçlar için.
Tek sarmallı RNA'da bulunan bilgi, polimer zincirinin spesifik bir purin ve pirimidin baz dizisi (yani birincil yapıda) formunda uygulanır. Bu dizi, RNA'nın "okunduğu" genin kodlama zincirini tamamlayıcı niteliktedir. Tamamlayıcılık nedeniyle, RNA molekülü, kodlayıcı zincire spesifik olarak bağlanabilir (melezleşebilir), ancak kodlamayan DNA zincirine hibritlenemez. RNA dizisi (T'nin U ile değiştirilmesi hariç), genin kodlamayan ipliğinin dizisiyle aynıdır (Şekil 37.8).
RNA'nın biyolojik fonksiyonları
Çeşitli RNA türleri bilinmektedir. Hemen hemen hepsi protein biyosentezi sürecine doğrudan katılmaktadır. Protein sentezi için şablon görevi gören sitoplazmik RNA moleküllerine haberci RNA (mRNA) adı verilir. Sitoplazmik RNA'nın başka bir türü olan ribozomal RNA (rRNA), ribozomların (oynayan organeller) yapısal bileşenleri olarak görev yapar. önemli rol protein sentezinde). Transfer RNA'sının (tRNA) adaptör molekülleri, mRNA bilgilerinin proteinlerdeki amino asit dizisine çevrilmesinde (translasyonunda) rol oynar.
Memeli hücreleri de dahil olmak üzere ökaryotik hücrelerde üretilen birincil RNA transkriptlerinin önemli bir kısmı çekirdekte bozunmaya uğrar ve sitoplazmada herhangi bir yapısal veya bilgilendirici rol oynamaz. Ekili olarak

Pirinç. 37.7. Bir RNA molekülünün ikincil yapısı, tamamlayıcı nükleik baz çiftleri arasındaki hidrojen bağlarının molekül içi oluşumundan kaynaklanan "kök ilmek" ("saç tokası") tipindedir.
İnsan hücrelerinde, protein sentezinde doğrudan yer almayan, ancak RNA işlemeyi ve hücrenin genel "mimarisini" etkileyebilen küçük nükleer RNA'lardan oluşan bir sınıf keşfedildi. Bu nispeten küçük moleküllerin boyutları değişiklik gösterir; ikincisi 90 ila 300 nükleotid içerir (Tablo 37.3).
RNA, bazı hayvan ve bitki virüslerinin ana genetik materyalidir. Bazı RNA virüsleri hiçbir zaman RNA'nın DNA'ya ters transkripsiyonuna uğramaz. Bununla birlikte, retrovirüsler gibi bilinen hayvan virüslerinin çoğu, çift sarmallı bir DNA kopyası oluşturmak üzere RNA'ya bağımlı DNA polimeraz (ters transkriptaz) tarafından yönlendirilen RNA genomlarının ters transkripsiyonu ile karakterize edilir. Çoğu durumda, ortaya çıkan çift sarmallı DNA transkript, genoma entegre edilir ve ardından viral genlerin ekspresyonunu ve ayrıca viral RNA genomlarının yeni kopyalarının üretimini sağlar.
RNA'nın yapısal organizasyonu
Tüm ökaryotik ve prokaryotik organizmalarda üç ana RNA molekülü sınıfı vardır: haberci RNA (mRNA), transfer RNA (tRNA) ve ribozomal RNA (rRNA). Bu sınıfların temsilcileri büyüklük, işlev ve stabilite açısından birbirinden farklıdır.
Messenger RNA (mRNA), boyut ve stabilite açısından en heterojen sınıftır. Bu sınıfın tüm temsilcileri, genden hücrenin protein sentezleme sistemine kadar bilgi taşıyıcısı olarak görev yapar. Sentezlenen polipeptit için şablon görevi görürler, yani. proteinin amino asit dizisini belirlerler (Şekil 37.9).
Haberci RNA'lar, özellikle ökaryotik olanlar, bazı benzersiz yapısal özelliklere sahiptir. MRNA'nın 5-ucu, bir trifosfat kalıntısı yoluyla bitişik 2-0-metilribonükleosidin 5-hidroksiline bağlanan bir 7-metilguanozin trifosfatla kapatılır (Şekil 37.10). mRNA molekülleri sıklıkla dahili 6-metiladenin kalıntıları ve 2-0-metillenmiş ribonükleotidler içerir. Her ne kadar "kapama"nın anlamı henüz tam olarak anlaşılmamış olsa da, mRNA'nın 5-ucunda ortaya çıkan yapının çeviri sisteminde spesifik tanıma için kullanıldığı varsayılabilir. Protein sentezi mRNA'nın 5" (başlıklı) ucunda başlar. Çoğu mRNA molekülünün diğer ucu (3-uç), 20-250 nükleotidlik bir poliadenilat zinciri içerir. Bunun spesifik işlevleri tam olarak belirlenmemiştir. bu yapının hücre içi mRNA stabilitesinin korunmasından sorumlu olduğu varsayılabilir. Histon mRNA'lar da dahil olmak üzere bazı mRNA'lar, poli (A) kuyruğunun hareketsizleştirilmiş oligo (T) ile tamamlayıcı etkileşimleri nedeniyle oluşan sütunla poli (A) içermez. ).
Pirinç. 37.8. Gen dizisi ve RNA transkripti. Kodlayan ve kodlamayan şeritler gösterilir ve polariteleri not edilir. Polariteye sahip bir RNA transkripti, kodlama zincirini tamamlayıcıdır (polaritesi 3 - 5 olan) ve kodlamayan DNA zincirinin sekansı (T'den U'ya ikameler hariç) ve polaritesi bakımından aynıdır.

Pirinç. 37.9. DNA genetik bilgisinin bir mRNA transkript formunda ekspresyonu ve ardından spesifik bir protein molekülünün oluşumu ile ribozomların katılımıyla translasyon.
(bkz: tarama)
Pirinç. 37.10. Çoğu ökaryotik haberci RNA'nın 5-ucunda bulunan "başlık" yapısı 7-metilguanozin trifosfat, mRNA'nın 5-ucuna bağlanır. genellikle bir 2-O-metilpurin nükleotidi içerir.
İnsan hücreleri de dahil olmak üzere memeli hücrelerinde, sitoplazmada bulunan olgun mRNA molekülleri, genin kopyalanan bölgesinin tam bir kopyası değildir. Transkripsiyon sonucu oluşan poliribonükleotid, sitoplazmik mRNA'nın öncüsüdür; çekirdekten ayrılmadan önce spesifik işlemlere tabi tutulur. Memeli hücrelerinin çekirdeklerinde bulunan tam uzunluktaki transkripsiyon ürünleri, dördüncü sınıf RNA moleküllerini oluşturur. Bu tür nükleer RNA'lar çok heterojendir ve önemli boyutlara ulaşır. Heterojen nükleer RNA molekülleri, mRNA'nın moleküler ağırlığı genellikle 2106'yı aşmazken, moleküler ağırlığa sahip olabilir. Çekirdekte işlenirler ve ortaya çıkan olgun mRNA'lar, protein biyosentezi için bir şablon görevi gördükleri sitoplazmaya girerler. .
Transfer RNA (tRNA) molekülleri tipik olarak yaklaşık 75 nükleotit içerir. Bu tür moleküllerin moleküler ağırlığı . tRNA'lar ayrıca karşılık gelen öncü moleküllerin spesifik işlenmesinin bir sonucu olarak da oluşturulur (bkz. Bölüm 39). Transfer tRNA'ları, mRNA'nın çevirisi sırasında aracı görevi görür. Herhangi bir hücrede en az 20 çeşit tRNA molekülü vardır. Her bir tRNA türü (bazen birkaç tür), protein sentezi için gerekli olan 20 amino asitten birine karşılık gelir. Her spesifik tRNA, nükleotid sekansı açısından diğerlerinden farklı olmasına rağmen, hepsinde ortak özellikler. Birkaç iplik içi tamamlayıcı bölge nedeniyle, tüm tRNA'lar "yonca yaprağı" adı verilen ikincil bir yapıya sahiptir (Şekil 37.11).
Tüm tRNA molekül türlerinin dört ana kolu vardır. Alıcı kol, eşleştirilmiş nükleotidlerden oluşan bir "gövde"den oluşur ve CCA dizisi ile biter. Amino asidin karboksil grubuna bağlanma, adenosil kalıntısının Y-hidroksil grubu aracılığıyla gerçekleşir. Geri kalan kollar ayrıca tamamlayıcı baz çiftleri ve eşleşmemiş bazların ilmeklerinden oluşan "gövdelerden" oluşur (Şekil 37.7). Antikodon kolu, mRNA'daki bir nükleotid üçlüsünü veya kodonunu (bkz. Bölüm 40) tanır. D kolu, içindeki dihidroüridin varlığı nedeniyle bu şekilde adlandırılmıştır; -kol, T-psödoüridin-C dizisinden sonra adlandırılmıştır. Aksesuar kolu en değişken yapıdır ve tRNA sınıflandırmasının temelini oluşturur. Sınıf 1 tRNA'ların (toplam sayılarının %75'i) 3-5 baz çifti uzunluğunda ek bir kolu vardır. Sınıf 2 tRNA moleküllerinin ekstra kolu 13-21 baz çiftinden oluşur ve sıklıkla eşlenmemiş bir döngü içerir.

Pirinç. 37.11. 3-CCA ucuna bir amino asit eklenmiş bir aminoasil-tRNA molekülünün yapısı. Molekül içi hidrojen bağları ve antikodon, TTC ve dihidrourasil kollarının konumu belirtilmiştir. (J.D. Watson'dan. Moleküler biyoloji of the Gene 3rd, ed.. Telif hakkı 1976, 1970, 1965, W.A. Benjamin, Inc., Menlo Park Calif.)
Karşılık gelen kolların nükleotid bazlarının tamamlayıcı etkileşimleri sistemi tarafından belirlenen ikincil yapı, tüm türlerin karakteristiğidir. Alıcı kol yedi baz çifti içerir, -kol beş baz çifti içerir ve D kolu üç (veya) içerir. dört) baz çiftleri.
tRNA molekülleri prokaryotlarda çok kararlıdır ve ökaryotlarda biraz daha az kararlıdır. Bunun tersi durum, prokaryotlarda oldukça kararsız olan ancak ökaryotik organizmalarda önemli bir stabiliteye sahip olan mRNA için tipiktir.
Ribozomal RNA. Bir ribozom, bir mRNA şablonu kullanılarak protein sentezi için tasarlanmış sitoplazmik bir nükleoprotein yapısıdır. Ribozom spesifik bir temas sağlar ve bunun sonucunda spesifik bir genden okunan nükleotid sekansı, karşılık gelen proteinin amino asit sekansına çevrilir.
Tabloda Şekil 37.2, 4.2106 moleküler ağırlığa ve sedimantasyon hızına (Svedberg birimleri) sahip memeli ribozomlarının bileşenlerini göstermektedir. Memeli ribozomları iki nükleoprotein alt biriminden oluşur; bunlardan büyük olanı
Tablo 37.2. Memeli ribozomlarının bileşenleri

moleküler ağırlık (60S) ve düşük moleküler ağırlık (40S). 608 alt birimi 58-ribozomal RNA (rRNA), 5.8S-pRNA ve 28S-pRNA'nın yanı sıra 50'den fazla farklı polipeptit içerir. Küçük 408 alt birimi, tek bir 18S-pRNA ve yaklaşık 30 polipeptit zinciri içerir. 5S-RNA dışındaki tüm ribozomal RNA'lar, nükleolusta lokalize olan ortak bir öncüye, 45S-RNA'ya sahiptir (bkz. Bölüm 40). 5S-RNA molekülünün kendi öncüsü vardır. Nükleolusta yüksek oranda metillenmiş ribozomal RNA'lar, ribozomal proteinlerle paketlenir. Sitoplazmada ribozomlar oldukça stabildir ve görevleri yerine getirme yeteneğine sahiptir. büyük sayı yayın döngüleri.
Küçük kararlı RNA'lar. Ökaryotik hücrelerde çok sayıda ayrı, yüksek oranda korunmuş, küçük ve kararlı RNA molekülleri bulunmuştur. Bu tipteki RNA'ların çoğu ribonükleoproteinlerde bulunur ve çekirdekte, sitoplazmada veya her iki bölmede aynı anda lokalize olur. Bu moleküllerin boyutları 90 ila 300 nükleotid arasında değişmekte olup içerikleri hücre başına 100.000-1.000.000 kopyadır.
Küçük nükleer ribonükleik parçacıklar (genellikle snurps olarak adlandırılır) muhtemelen gen ifadesinin düzenlenmesinde önemli bir rol oynar. U7 tipi nükleoprotein parçacıklarının, histon mRNA'larının 3-terminallerinin oluşumunda rol oynadığı görülmektedir. Parçacıklar muhtemelen poliadenilasyon, a - intronların uzaklaştırılması ve mRNA işlenmesi için gereklidir (bkz. Bölüm 39). Masa 37.3. küçük kararlı RNA'ların bazı özelliklerini özetler.
Tablo 37.3. Memeli hücrelerinde bulunan bazı küçük kararlı RNA türleri

EDEBİYAT
Darnell J. ve ark. Moleküler Hücre Biyolojisi, Scientific American Books, 1986.
Hunt T. DNA, RNA'yı Protein Yapar, Elsevier, 1983. Lewin B. Genes, 2. baskı, Wiley, 1985.
Zengin A. ve ark. Solak Z-DNA'nın kimyası ve biyolojisi, Annu. Rev. Biochem., 1984, 53, 847.
Turner P. Snurps için kontrol rolleri, Nature, 1985, 316, 105. Watson J. D. The Double Helix, Atheneum, 1968.
Watson J.D., Crick F.H.C. Nükleik asitlerin moleküler yapısı. Doğa, 1953, 171, 737.
Zieve G. W. İki grup küçük stabil RNA, Cell, 1981, 25, 296.
Biyolojik Bilimler Adayı S. GRIGOROVICH.
Tarihin ilk şafağında, insan aklı ve bununla birlikte soyut düşünme yeteneğini kazandığında, her şeyi açıklamaya yönelik karşı konulamaz bir ihtiyacın esiri oldu. Güneş ve Ay neden parlıyor? Nehirler neden akar? Dünya nasıl çalışıyor? Elbette en önemlilerinden biri yaşamın özü sorusuydu. Yaşayanlar ile büyüyenler ile ölüler ve hareketsizler arasındaki keskin fark göz ardı edilemeyecek kadar çarpıcıydı.
D. Ivanovsky tarafından 1892'de tanımlanan ilk virüs, tütün mozaik virüsüydü. Bu keşif sayesinde hücreden daha ilkel canlıların olduğu ortaya çıktı.
Rus mikrobiyolog D. I. Ivanovsky (1864-1920), virolojinin kurucusu.
1924 yılında A. I. Oparin (1894-1980), genç Dünya'nın hidrojen, metan, amonyak, karbondioksit ve su buharından oluşan atmosferinde amino asitlerin sentezlenebileceğini ve bunların daha sonra kendiliğinden proteinlere birleştirilebileceğini öne sürdü.
Amerikalı biyolog Oswald Avery, bakterilerle yapılan deneylerde kalıtsal özelliklerin aktarılmasından sorumlu olanın nükleik asitler olduğunu ikna edici bir şekilde gösterdi.
RNA ve DNA'nın karşılaştırmalı yapısı.
İki boyutlu mekansal yapı Tek hücreli organizma Tetrahymena'nın ribozimi.
Protein sentezi için moleküler bir makine olan ribozomun şematik gösterimi.
“İn vitro evrim” sürecinin şeması (Selex yöntemi).
Louis Pasteur (1822-1895), aynı maddenin (tartarik asit) kristallerinin iki ayna simetrik uzaysal konfigürasyona sahip olabileceğini keşfeden ilk kişiydi.
1950'lerin başında, Chicago Üniversitesi'nden (ABD) Stanley Miller, simülasyonu simüle eden ilk deneyi gerçekleştirdi. kimyasal reaksiyonlar Genç Dünya'nın koşullarında meydana gelebilecek olan.
Amino asitler gibi kiral moleküller, solak ve sol yönlü gibi ayna simetriktir. sağ el. "Kiralite" teriminin kendisi Yunanca "chiros" - el kelimesinden gelir.
RNA dünyasının teorisi.
Bilim ve yaşam // İllüstrasyonlar
Tarihin her aşamasında insanlar, gezegenimizde yaşamın ortaya çıkışı bilmecesine çözümlerini sundular. "Bilim" kelimesini bilmeyen eskiler, bilinmeyene basit ve anlaşılır bir açıklama buldular: "Etrafındaki her şey bir zamanlar biri tarafından yaratılmıştı." Tanrılar böyle ortaya çıktı.
Mısır'da, Çin'de ve daha sonra beşikte eski uygarlıkların doğuşundan bu yana modern bilim- Yunanistan'da Orta Çağ'a kadar dünyayı anlamanın temel yöntemi "otoritelerin" gözlemleri ve görüşleriydi. Sürekli gözlemler, canlıların belirli koşullar altında cansızlardan ortaya çıktığını açıkça gösterdi: sivrisinekler ve timsahlar bataklık çamurundan, sinekler çürüyen yiyeceklerden ve fareler üzerine buğday serpilmiş kirli çamaşırlardan. Sadece belirli bir sıcaklık ve nemi korumak önemlidir.

Dünyanın yaratılışına ve ilahi planların anlaşılmazlığına ilişkin dini dogmaya dayanan Orta Çağ Avrupalı "bilim adamları", yaşamın kökeni hakkında yalnızca İncil ve dini yazılar çerçevesinde tartışmanın mümkün olduğunu düşünüyorlardı. Allah'ın yarattığı şeylerin özü anlaşılamaz, ancak kutsal metinlerdeki bilgilerle veya ilahi ilhamın etkisiyle "açıklanabilir". O zamanlar hipotezleri test etmenin kötü bir biçim olduğu düşünülüyordu ve Kutsal Kilise'nin görüşünü sorgulamaya yönelik herhangi bir girişim, hoş olmayan bir mesele, sapkınlık ve saygısızlık olarak görülüyordu.
Yaşam bilgisi zamanı işaretliyordu. Filozofların başarıları iki bin yıl boyunca bilimsel düşüncenin zirvesi olarak kaldı Antik Yunanistan. Bunların en önemlileri Platon (MÖ 428/427 - 347) ve öğrencisi Aristoteles'tir (MÖ 384 - 322). Platon, diğer şeylerin yanı sıra, başlangıçta cansız olan maddeye ölümsüz, maddi olmayan bir ruhun - "psişenin" - aşılanması yoluyla canlandırılması fikrini önerdi. Canlıların cansızlardan kendiliğinden türediği teorisi böyle ortaya çıktı.
Bilim için harika bir sözcük olan "deney" Rönesans'la birlikte ortaya çıktı. Bir kişinin eski bilim adamlarının yetkili açıklamalarının değişmezliğinden şüphe etmeye karar vermesi iki bin yıl sürdü. Bildiğimiz ilk cesurlardan biri İtalyan doktor Francisco Redi'ydi (1626 - 1698). Son derece basit ama etkili bir deney gerçekleştirdi: Birkaç kaba bir parça et yerleştirerek bazılarını kapladı. kalın kumaş, diğerleri gazlı bezle ve diğerleri açık bırakıldı. Sinek larvalarının yalnızca açık kaplarda (sineklerin konabileceği) gelişmesi, ancak kapalı kaplarda (hala havaya erişimi olan) gelişmemesi, Platon ve Aristoteles'in destekçilerinin anlaşılmaz hakkındaki inançlarıyla keskin bir şekilde çelişiyordu. canlılık havada süzülüyor ve cansız maddeyi canlı maddeye dönüştürüyor.
Bu ve benzeri deneyler, iki grup bilim insanı arasında şiddetli savaşların başladığı bir dönemin başlangıcına işaret ediyordu: vitalistler ve mekanistler. Tartışmanın özü şu soruydu: "Canlıların işleyişi (ve görünümü), cansız maddeler için de geçerli olan fizik kanunlarıyla açıklanabilir mi?" Vitalistler olumsuz yanıt verdi. “Hücre ancak hücredendir, yaşayan her şey ancak canlıdandır!” 19. yüzyılın ortalarında ortaya atılan bu konum, vitalizmin bayrağı haline geldi. Bu tartışmadaki en paradoksal şey, bugün bile, vücudumuzu oluşturan atom ve moleküllerin "cansız" doğasını bilen ve genel olarak mekanik bakış açısına katılan bilim adamlarının, bu olayın olasılığı konusunda deneysel bir doğrulamaya sahip olmamasıdır. cansız maddeden hücresel yaşamın ortaya çıkışı. Henüz hiç kimse, canlı organizmaların dışında bulunan "inorganik" "parçalardan" en ilkel hücreyi bile "oluşturmayı" başaramamıştır. Bu, çığır açan bu tartışmada son noktanın henüz belirlenmediği anlamına geliyor.
Peki yaşam Dünya'da nasıl ortaya çıkabilir? Mekanistlerin konumlarını paylaşarak, elbette, yaşamın ilk olarak çok basit, ilkel bir şekilde ortaya çıkması gerektiğini hayal etmek en kolay yoldur. düzenlenmiş form. Ancak yapının basitliğine rağmen yine de Hayat olmalı, yani canlıyı cansızdan ayıran asgari özelliklere sahip bir şey.
Yaşam için kritik olan bu özellikler nelerdir? Yaşamayı, cansızdan ayıran şey tam olarak nedir?
İle XIX sonu Yüzyıllar boyunca bilim adamları, tüm canlıların hücrelerden oluştuğuna ikna olmuşlardı ve bu, onlarla cansız maddeler arasındaki en belirgin farktır. Bilinen tüm hücrelerden daha küçük olmasına rağmen diğer organizmaları aktif olarak enfekte edebilen, içlerinde çoğalabilen ve aynı (veya çok benzer) biyolojik özelliklere sahip yavrular üretebilen virüslerin keşfine kadar buna inanılıyordu. Keşfedilen ilk virüs olan tütün mozaik virüsü, 1892 yılında Rus bilim adamı Dmitry Ivanovsky (1864-1920) tarafından tanımlandı. O zamandan bu yana, hücrelerden daha ilkel canlıların da Yaşam olarak adlandırılma hakkına sahip olabileceği anlaşıldı.
Virüslerin ve daha sonra canlıların daha ilkel biçimlerinin - viroidlerin keşfi, sonuçta, incelenen nesnenin canlı olarak adlandırılması için gerekli ve yeterli olan minimum bir dizi özelliği formüle etmeyi mümkün kıldı. Öncelikle kendi türünü yeniden üretebilme yeteneğine sahip olması gerekir. Ancak bu tek şart değildir. Varsayımsal bir ilkel yaşam maddesi (örneğin, ilkel bir hücre veya molekül) basitçe kendi kendisini üretebilseydi tam kopyalar sonuçta değişen koşullarda hayatta kalamayacaktı çevre genç bir Dünya'da daha karmaşık formların oluşması (evrim) imkansız hale gelirdi. Sonuç olarak, sözde ilkel "ilkel yaşamın özü", olabildiğince basit tasarlanmış, ancak aynı zamanda özelliklerini değiştirebilen ve sonraki nesillere aktarabilen bir şey olarak tanımlanabilir.
RNA genellikle bir sarmal şeklinde bükülmüş bir iplikçikten oluşur. Virüsler çift sarmallı RNA'ya sahiptir. RNA, nükleolus, çekirdek, sitoplazma ve ribozomlarda bulunur. RNA molekülleri DNA moleküllerinden daha kısadır.
RNA türleri
Üç tür RNA vardır: ribozomal, matris (haberci - mRNA), taşıma (tRNA). Hücredeki konumları, boyutları, nükleotid bileşimleri ve fonksiyonel özellikleri bakımından birbirlerinden farklılık gösterirler.
RNA, enzimlerin katılımıyla sentezlenir RNA polimerazları bir DNA molekülü üzerinde. Bir DNA molekülünün bir bölümünün nükleotid dizisi, RNA molekülündeki nükleotidlerin sırasını belirler.
Çoğu hücre, DNA içeriğinden önemli ölçüde daha yüksek RNA içeriğine (5 ila 10 kat) sahiptir. En büyük kısım RNA ribozomaldir.
RNA'nın fonksiyonları
RNA'nın fonksiyonları: kalıtsal bilgiyi uygular, protein sentezinde yer alır.
Bilgi(matris) RNA (mRNA), DNA'nın bir bölümünün, yani bir veya daha fazla genin kopyasıdır. Genetik bilgiyi polipeptit zincirinin sentez bölgesine aktarır ve doğrudan buna dahil olur. RNA'nın kopyaladığı DNA bölümünün uzunluğuna göre 300-30.000 nükleotidden oluşur. Hücredeki kısım ve RNA toplamın yaklaşık %5'ini oluşturur. Moleküller ve RNA nispeten kararsızdır; hızla nükleotidlere ayrılırlar. Ökaryotik hücrelerde ömürleri birkaç saate kadar, mikroorganizmalarda ise birkaç dakikaya kadardır.
DNA molekülü gibi RNA'nın da hidrojen bağları, hidrofobik, elektrostatik etkileşimler vb. yoluyla oluşturulan ikincil ve üçüncül yapıları vardır.
ribozomal RNA, ribozom kütlesinin %60'ını, yani hücredeki toplam RNA miktarının yaklaşık %85'ini oluşturur. 3000-5000 nükleotid içerir. Kalıtsal bilgilerin aktarımında yer almaz. Ribozomun bir parçasıdır ve ökaryotlarda yaklaşık 100 tane bulunan proteinleriyle etkileşime girer. Ökaryotlarda dört tür ribozomal RNA bulunur, prokaryotlarda ise üç tür bulunur. Yapısal bir işlevi yerine getirir: ribozom üzerinde mRNA ve tRNA'nın spesifik bir mekansal düzenlemesini sağlar.
Taşıma (tRNA) - amino asitleri protein sentezi bölgesine taşır. Tamamlayıcılık ilkesine dayanarak mRNA'nın taşınan amino asite karşılık gelen bölgesini tanır. Her amino asit, protein sentezi bölgesine kendi tRNA'sı tarafından taşınır. tRNA'lar hücre hücre iskeletinin elemanları tarafından taşınır.
Hidrojen bağları tarafından sağlanan kalıcı bir ikincil yapı olan yonca yaprağı (yonca yaprağı) şeklindedir. TRNA'nın tepesinde mRNA kodonuna karşılık gelen ve adı verilen bir üçlü nükleotid vardır. antikodon . Bazın yakınında kovalent bağ sayesinde amino asit molekülünün bağlandığı bir bölge vardır. tRNA 70-90 nükleotid içerir. Toplam RNA miktarının %10'unu oluşturur. Yaklaşık 60 çeşit tRNA bilinmektedir.
tRNA oldukça kompakt L benzeri bir yapıya sahip olabilir düzensiz şekil üçüncül yapı.
Dinükleotidler
İki nükleotidden oluşurlar ancak yapısal özelliklere sahiptirler. En ünlüleri şunlardır: nikotinamid adenin dinükleotid (NAD +), nikotinamid adenin dinükleotid fosfat (NADP +). Ana fonksiyon elektronların (2) ve hidrojen iyonlarının (1) transferidir. Geri yüklenebilir:
NAD + + 2e - + H + →NADH;
NADP + + 2e - + H + →NADPH.
Bazı reaksiyonlar için belirli bir bölgede bu bileşikler bir hidrojen protonu ve elektronları bağışlar:
NADH →NAD + + 2e - + H +;
NADPH → NADP + + 2e - + H +
Polinükleotidin yapısal biriminde hangi monosakkaritin bulunduğuna bağlı olarak - riboz veya 2-deoksiriboz, ayırt etmek
- ribonükleik asitler (RNA) ve
- deoksiribonükleik asitler(DNA).
DNA makromoleküllerinin nükleotid birimleri şunları içerebilir: adenin, guanin, sitozin Ve timin. RNA'nın bileşimi şu şekilde farklılık gösterir: Timina Sunmak urasil.
DNA'nın moleküler ağırlığı on milyonlarca amu'ya ulaşır. Bunlar bilinen en uzun makromoleküllerdir. RNA'nın moleküler ağırlığı önemli ölçüde daha düşüktür (birkaç yüzden onbinlere kadar). DNA esas olarak hücrelerin çekirdeğinde, RNA ise ribozomlarda ve hücrelerin protoplazmasında bulunur.
Nükleik asitlerin yapısını açıklarken dikkate alın farklı seviyeler makromoleküllerin organizasyonu: öncelik Ve ikincil yapı.
- Birincil yapı nükleik asitler bu, nükleotit bileşimi ve polimer zincirindeki belirli bir nükleotit birimi dizisidir.
Kısaltılmış tek harfli gösterimde bu yapı şu şekilde yazılır: ...– A – G – C –...
- Altında ikincil yapı Nükleik asitler, polinükleotid zincirlerinin uzaysal olarak sıralanmış formlarını anlar.

Bu uzaysal yapı, sarmal içine yönlendirilen nitrojenli bazların oluşturduğu birçok hidrojen bağı tarafından yerinde tutulur. Bir zincirin purin bazı ile diğer zincirin pirimidin bazı arasında hidrojen bağları oluşur. Bu bazlar tamamlayıcı çiftler oluşturur (enlem. tamamlayıcı- ek). Tamamlayıcı baz çiftleri arasında hidrojen bağlarının oluşumu bunların uzaysal yazışmalarından kaynaklanmaktadır. Bir pirimidin bazı, bir pürin bazının tamamlayıcısıdır:

Diğer baz çiftleri arasındaki hidrojen bağları, bunların çift sarmal yapıya yerleşmesini engeller. Böylece,
- TİMİN (T), ADENİN (A)'nın tamamlayıcısıdır,
- SİTOSİN (C), GUANİN'in (G) tamamlayıcısıdır.

Polinükleotid zincirlerinin tamamlayıcılığı, DNA'nın ana işlevi olan kalıtsal özelliklerin depolanması ve iletilmesi için kimyasal temel görevi görür.
DNA'nın yalnızca genetik bilgiyi saklama değil aynı zamanda kullanma yeteneği de aşağıdaki özellikleriyle belirlenir:
RNA'nın ikincil yapısı. DNA'nın aksine, RNA molekülleri tek bir polinükleotid zincirinden oluşur ve kesin olarak tanımlanmış bir uzaysal şekle sahip değildir (RNA'nın ikincil yapısı biyolojik işlevlerine bağlıdır).
RNA'nın ana rolü protein biyosentezine doğrudan katılımdır. Hücredeki konumu, bileşimi, boyutu ve protein makromoleküllerinin oluşumundaki spesifik rolünü belirleyen özellikleri bakımından farklılık gösteren üç tip hücresel RNA bilinmektedir:
- Haberci RNA'lar, DNA'da kodlanan proteinin yapısı hakkındaki bilgileri hücre çekirdeğinden, protein sentezinin gerçekleştiği ribozomlara iletir;
- transfer RNA'ları hücre sitoplazmasındaki amino asitleri toplar ve bunları ribozoma aktarır; Bu tip RNA molekülleri, protein sentezine hangi amino asitlerin katılması gerektiğini haberci RNA zincirinin karşılık gelen bölümlerinden "öğrenir";
- Ribozomal RNA'lar, haberci RNA'dan gelen bilgileri okuyarak belirli bir yapıdaki proteinin sentezini sağlar.



