Genetik kod, bir proteindeki amino asitlere karşılık gelen kodonları oluşturan DNA veya RNA'daki belirli bir nükleotit dizilim değişikliğine dayalı olarak nükleik asit moleküllerindeki kalıtsal bilgileri kaydetmek için bir sistemdir.
Genetik kodun özellikleri.
Genetik kodun çeşitli özellikleri vardır.
üçlülük.
Dejenerasyon veya fazlalık.
belirsizlik.
Polarite.
Örtüşmeyen.
Kompaktlık.
çok yönlülük
Bazı yazarların, koda dahil edilen nükleotitlerin kimyasal özellikleri veya vücudun proteinlerinde tek tek amino asitlerin oluşma sıklığı vb. ile ilgili kodun başka özelliklerini de sunduğuna dikkat edilmelidir. Ancak, bu özellikler yukarıdan takip eder, bu yüzden onları orada ele alacağız.
a. üçlülük. Genetik kod, birçok karmaşık şekilde organize edilmiş sistem gibi, en küçük yapısal ve en küçük işlevsel birime sahiptir. Üçlü, genetik kodun en küçük yapısal birimidir. Üç nükleotitten oluşur. Bir kodon, genetik kodun en küçük işlevsel birimidir. Kural olarak, mRNA üçlülerine kodon denir. Genetik kodda, bir kodon birkaç işlevi yerine getirir. İlk olarak, ana işlevi, bir amino asidi kodlamasıdır. İkincisi, bir kodon bir amino asidi kodlamayabilir, ancak bu durumda farklı bir işlevi vardır (aşağıya bakınız). Tanımdan da anlaşılacağı gibi, üçlü karakterize eden bir kavramdır. temel yapısal birim genetik kod (üç nükleotid). kodon karakterize eder temel anlam birimi genom - üç nükleotit, bir amino asidin polipeptit zincirine bağlanmayı belirler.
Temel yapı birimi önce teorik olarak deşifre edildi, ardından varlığı deneysel olarak doğrulandı. Gerçekten de 20 amino asit bir veya iki nükleotit tarafından kodlanamaz. ikincisi sadece 4'tür. Dört nükleotitten üçü, canlı organizmalarda bulunan amino asitlerin sayısını fazlasıyla karşılayan 4 3 = 64 varyant verir (bkz. Tablo 1).
Tablo 64'te sunulan nükleotid kombinasyonlarının iki özelliği vardır. İlk olarak, 64 üçlü varyanttan sadece 61'i kodondur ve herhangi bir amino asidi kodlar, bunlara denir. duyu kodonları. Üç üçlü kodlama yapmıyor
amino asitler a, çevirinin sonunu işaret eden durdurma sinyalleridir. Böyle üç üçüz var UAA, UAG, UGA, aynı zamanda "anlamsız" (saçma kodonlar) olarak da adlandırılırlar. Bir üçlüdeki bir nükleotidin bir başkasıyla yer değiştirmesi ile ilişkili bir mutasyon sonucunda, bir duyu kodonundan anlamsız bir kodon ortaya çıkabilir. Bu tür mutasyon denir anlamsız mutasyon. Genin içinde (bilgi kısmında) böyle bir durma sinyali oluşursa, bu yerdeki protein sentezi sırasında süreç sürekli olarak kesintiye uğrayacaktır - proteinin sadece ilk (durma sinyalinden önce) kısmı sentezlenecektir. Böyle bir patolojiye sahip bir kişi, protein eksikliği yaşayacak ve bu eksiklikle ilişkili semptomlar yaşayacaktır. Örneğin, hemoglobin beta zincirini kodlayan gende bu tür bir mutasyon bulundu. Hızla yok edilen kısaltılmış bir inaktif hemoglobin zinciri sentezlenir. Sonuç olarak, beta zincirinden yoksun bir hemoglobin molekülü oluşur. Böyle bir molekülün görevlerini tam olarak yerine getirmesinin olası olmadığı açıktır. Hemolitik aneminin tipine göre gelişen ciddi bir hastalık vardır (Yunanca "Talas" kelimesinden beta-sıfır talasemi - bu hastalığın ilk keşfedildiği Akdeniz).
Durdurma kodonlarının etki mekanizması, duyu kodonlarının etki mekanizmasından farklıdır. Bu, amino asitleri kodlayan tüm kodonlar için karşılık gelen tRNA'ların bulunması gerçeğinden kaynaklanmaktadır. Anlamsız kodonlar için hiçbir tRNA bulunamadı. Bu nedenle, tRNA, protein sentezini durdurma sürecinde yer almaz.
kodonAğustos (bazen bakterilerde GUG) sadece metiyonin ve valin amino asitini kodlamakla kalmaz, aynı zamandayayın başlatıcı .
b. Dejenerasyon veya fazlalık.
64 üçlüden 61'i 20 amino asidi kodlar. Üçlü sayısının amino asit sayısına göre üç kat fazla olması, bilgi transferinde iki kodlama seçeneğinin kullanılabileceğini düşündürmektedir. İlk olarak, 20 amino asidin kodlanmasında 64 kodonun tümü yer alamaz, ancak yalnızca 20 amino asit birkaç kodon tarafından kodlanabilir. Araştırmalar, doğanın ikinci seçeneği kullandığını göstermiştir.
Tercihi belli. 64 üçlüden sadece 20'si amino asitleri kodlamaya dahil olsaydı, 44 üçlü (64 üçlüden) kodlamaz kalır, yani. anlamsız (saçma kodonlar). Daha önce, mutasyonun bir sonucu olarak kodlama üçlüsünün anlamsız bir kodona dönüşmesinin hücrenin yaşamı için ne kadar tehlikeli olduğuna işaret etmiştik - bu, RNA polimerazın normal çalışmasını önemli ölçüde bozar ve sonuçta hastalıkların gelişmesine yol açar. Şu anda genomumuzda üç anlamsız kodon var ve şimdi anlamsız kodonların sayısı yaklaşık 15 kat artarsa ne olacağını hayal edin. Böyle bir durumda normal kodonların anlamsız kodonlara geçişinin ölçülemeyecek kadar yüksek olacağı açıktır.
Bir amino asidin birkaç üçlü tarafından kodlandığı bir koda dejenere veya fazlalık denir. Hemen hemen her amino asidin birkaç kodonu vardır. Böylece, amino asit lösin altı üçlü - UUA, UUG, CUU, CUC, CUA, CUG tarafından kodlanabilir. Valin dört üçlü, fenilalanin iki ve sadece üçlü ile kodlanır. triptofan ve metionin bir kodon tarafından kodlanmıştır. Aynı bilginin farklı karakterlerle kaydedilmesiyle ilişkilendirilen özelliğe denir. yozlaşma.
Bir amino aside atanan kodonların sayısı, amino asidin proteinlerde meydana gelme sıklığı ile iyi bir korelasyon içindedir.
Ve bu büyük olasılıkla tesadüfi değildir. Bir proteinde bir amino asidin ortaya çıkma sıklığı ne kadar yüksek olursa, bu amino asidin kodonu genomda ne kadar sık temsil edilirse, mutajenik faktörler tarafından zarar görme olasılığı o kadar yüksek olur. Bu nedenle, mutasyona uğramış bir kodonun, yüksek derecede dejenere olması durumunda aynı amino asidi kodlaması daha olasıdır. Bu konumlardan, genetik kodun yozlaşması, insan genomunu hasardan koruyan bir mekanizmadır.
Dejenerasyon teriminin moleküler genetikte başka bir anlamda da kullanıldığını belirtmek gerekir. Kodondaki bilgilerin ana kısmı ilk iki nükleotidin üzerine düştüğü için, kodonun üçüncü konumundaki bazın çok az önemli olduğu ortaya çıkıyor. Bu fenomene “üçüncü bazın yozlaşması” denir. İkinci özellik, mutasyonların etkisini en aza indirir. Örneğin kırmızı kan hücrelerinin temel işlevinin akciğerlerden dokulara oksijen, dokulardan da akciğerlere karbondioksit taşınması olduğu bilinmektedir. Bu işlev, eritrositin tüm sitoplazmasını dolduran solunum pigmenti - hemoglobin tarafından gerçekleştirilir. Karşılık gelen gen tarafından kodlanan bir protein parçası - globinden oluşur. Proteine ek olarak, hemoglobin demir içeren hem içerir. Globin genlerindeki mutasyonlar, farklı hemoglobin varyantlarının ortaya çıkmasına neden olur. Çoğu zaman, mutasyonlar aşağıdakilerle ilişkilidir: bir nükleotidin diğeriyle yer değiştirmesi ve gende yeni bir kodonun ortaya çıkması Hemoglobin polipeptit zincirinde yeni bir amino asidi kodlayabilen . Bir üçlüde, bir mutasyonun sonucu olarak, herhangi bir nükleotit değiştirilebilir - birinci, ikinci veya üçüncü. Birkaç yüz mutasyonun globin genlerinin bütünlüğünü etkilediği bilinmektedir. Yakın 400 bunların gendeki tek nükleotitlerin değiştirilmesi ve polipeptidde karşılık gelen amino asit ikamesi ile ilişkilidir. Bunlardan sadece 100 ikameler hemoglobin dengesizliğine ve hafiften çok şiddetliye kadar çeşitli hastalıklara yol açar. 300 (yaklaşık %64) sübstitüsyon mutasyonu hemoglobin fonksiyonunu etkilemez ve patolojiye yol açmaz. Bunun nedenlerinden biri, serin, lösin, prolin, arginin ve diğer bazı amino asitleri kodlayan üçlüdeki üçüncü nükleotidin yer değiştirmesi eşanlamlı bir kodonun ortaya çıkmasına neden olduğunda yukarıda bahsedilen “üçüncü bazın dejenerasyonu”dur. aynı amino asidi kodlayan Fenotipik olarak, böyle bir mutasyon kendini göstermeyecektir. Buna karşılık, vakaların %100'ünde bir üçlüdeki birinci veya ikinci nükleotidin herhangi bir şekilde değiştirilmesi, yeni bir hemoglobin varyantının ortaya çıkmasına neden olur. Ancak bu durumda bile ciddi fenotipik bozukluklar olmayabilir. Bunun nedeni, hemoglobindeki bir amino asidin fizikokimyasal özellikler açısından birincisine benzer bir amino asit ile yer değiştirmesidir. Örneğin, hidrofilik özelliklere sahip bir amino asit, aynı özelliklere sahip başka bir amino asit ile değiştirilirse.
Hemoglobin, bir demir porfirin grubundan (buna oksijen ve karbon dioksit molekülleri eklenir) ve bir protein - globinden oluşur. Yetişkin hemoglobin (HbA) iki özdeş içerir - zincirler ve iki -zincirler. molekül -zincir 141 amino asit kalıntısı içerir, - zincir - 146, - ve -zincirler birçok amino asit kalıntısında farklılık gösterir. Her globin zincirinin amino asit dizisi, kendi geni tarafından kodlanır. gen kodlama - zincir, kromozom 16'nın kısa kolunda bulunur, -gen - kromozom 11'in kısa kolunda. Gen kodlamasındaki değişiklik - birinci veya ikinci nükleotidin hemoglobin zinciri hemen hemen her zaman proteinde yeni amino asitlerin ortaya çıkmasına, hemoglobin fonksiyonlarının bozulmasına ve hasta için ciddi sonuçlara yol açar. Örneğin, CAU (histidin) üçlülerinden birinde “C” yi “U” ile değiştirmek, başka bir amino asit - tirozini kodlayan yeni bir UAU üçlüsünün ortaya çıkmasına neden olacaktır.Fenotipik olarak, bu ciddi bir hastalıkta kendini gösterecektir .. A 63. pozisyonda benzer değiştirme - histidin polipeptidinin tirozine zinciri, hemoglobini destabilize edecektir. Hastalık methemoglobinemi gelişir. 6. pozisyonda glutamik asidin valine mutasyon sonucu değişmesi zincir ciddi bir hastalığın nedenidir - orak hücreli anemi. Üzücü listeye devam etmeyelim. Sadece, ilk iki nükleotidi değiştirirken, bir amino asidin fizikokimyasal özelliklerde öncekine benzer görünebileceğini not ediyoruz. Böylece glutamik asidi (GAA) kodlayan üçlülerden birinde 2. nükleotidin yer değiştirmesi "Y" üzerindeki zincir, valini kodlayan yeni bir üçlünün (GUA) ortaya çıkmasına neden olur ve ilk nükleotidin "A" ile değiştirilmesi, amino asit lizini kodlayan bir AAA üçlüsü oluşturur. Glutamik asit ve lisin fizikokimyasal özelliklerde benzerdir - ikisi de hidrofiliktir. Valin hidrofobik bir amino asittir. Bu nedenle, hidrofilik glutamik asidin hidrofobik valin ile değiştirilmesi, hemoglobinin özelliklerini önemli ölçüde değiştirir, bu da sonuçta orak hücre anemisinin gelişmesine yol açarken, hidrofilik glutamik asidin hidrofilik lizin ile değiştirilmesi hemoglobinin işlevini daha az ölçüde değiştirir - hastalar hafif bir anemi formu geliştirin. Üçüncü bazın yer değiştirmesinin bir sonucu olarak, yeni üçlü, bir öncekiyle aynı amino asitleri kodlayabilir. Örneğin, CAH üçlüsünde urasil sitozin ile değiştirildiyse ve bir CAC üçlüsü ortaya çıktıysa, bir kişide pratik olarak hiçbir fenotipik değişiklik saptanmayacaktır. Bu anlaşılabilir, çünkü Her iki üçüz de aynı amino asit olan histidin'i kodlar.
Sonuç olarak, genetik kodun yozlaşmasının ve üçüncü bazın genel bir biyolojik konumdan yozlaşmasının, DNA ve RNA'nın benzersiz yapısında evrime dahil olan koruyucu mekanizmalar olduğunu vurgulamak uygundur.
içinde. belirsizlik.
Her üçlü (anlamsız olanlar hariç) sadece bir amino asidi kodlar. Böylece, kodon - amino asit yönünde, genetik kod belirsizdir, amino asit - kodon yönünde - belirsizdir (dejenere).
açık
kodon amino asit
dejenere
Ve bu durumda, genetik koddaki belirsizlik ihtiyacı açıktır. Başka bir varyantta, aynı kodonun translasyonu sırasında, protein zincirine farklı amino asitler eklenecek ve bunun sonucunda farklı birincil yapılara ve farklı işlevlere sahip proteinler oluşacaktır. Hücrenin metabolizması "bir gen - birkaç polipeptit" çalışma moduna geçecektir. Böyle bir durumda genlerin düzenleyici işlevinin tamamen kaybolacağı açıktır.
g. Polarite
DNA'dan ve mRNA'dan bilgi okumak sadece bir yönde gerçekleşir. Polarite, daha yüksek dereceli yapıları (ikincil, üçüncül, vb.) tanımlamak için gereklidir. Daha önce, daha düşük düzeydeki yapıların daha yüksek düzeydeki yapıları belirlediği gerçeğinden bahsetmiştik. Sentezlenen RNA zinciri DNA molekülünden uzaklaşır veya polipeptit zinciri ribozomdan uzaklaşır uzaklaşmaz, proteinlerde üçüncül yapı ve daha yüksek bir düzenin yapıları hemen oluşur. RNA veya polipeptidin serbest ucu üçüncül bir yapı kazanırken, zincirin diğer ucu DNA (eğer RNA kopyalanmışsa) veya ribozom (polipeptit kopyalanmışsa) üzerinde sentezlenmeye devam eder.
Bu nedenle, tek yönlü bilgi okuma işlemi (RNA ve protein sentezinde) yalnızca sentezlenen maddedeki nükleotitlerin veya amino asitlerin sırasını belirlemek için değil, aynı zamanda ikincil, üçüncül vb. yapılar.
e. Örtüşmeyen.
Kod çakışabilir veya çakışmayabilir. Çoğu organizmada, kod örtüşmez. Bazı fajlarda örtüşen bir kod bulundu.
Örtüşmeyen bir kodun özü, bir kodonun nükleotidinin aynı anda başka bir kodonun nükleotidi olamamasıdır. Kod örtüşüyorsa, yedi nükleotid dizisi (GCUCUG) örtüşmeyen bir kod durumunda olduğu gibi iki amino asidi (alanin-alanin) (Şekil 33, A) değil, üçünü (eğer bir nükleotid ise) kodlayabilirdi. yaygındır) (Şekil 33, B) veya beş (eğer iki nükleotid ortaksa) (bkz. Şekil 33, C). Son iki durumda, herhangi bir nükleotidin mutasyonu, iki, üç vb. dizide bir ihlale yol açacaktır. amino asitler.
Bununla birlikte, bir nükleotidin mutasyonunun, bir polipeptite bir amino asidin dahil edilmesini her zaman bozduğu bulunmuştur. Bu, kodun örtüşmediği gerçeği lehinde önemli bir argümandır.
Bunu Şekil 34'te açıklayalım. Kalın çizgiler, örtüşmeyen ve örtüşen kod durumunda amino asitleri kodlayan üçlüleri göstermektedir. Deneyler, genetik kodun örtüşmediğini açıkça göstermiştir. Deneyin detaylarına girmeden, nükleotid dizisindeki üçüncü nükleotidi değiştirirsek (bkz. Şekil 34) not ediyoruz.saat (yıldızla işaretlenmiştir) o zaman bir başkasına:
1. Örtüşmeyen bir kodla, bu dizi tarafından kontrol edilen protein, bir (birinci) amino asit (yıldızla işaretlenmiş) için bir yedeğine sahip olacaktır.
2. A seçeneğinde çakışan bir kodla, iki (birinci ve ikinci) amino asitte (yıldızla işaretlenmiş) bir yer değiştirme gerçekleşir. B seçeneği altında, ikame üç amino asidi etkileyecektir (yıldızlarla işaretlenmiştir).
Bununla birlikte, çok sayıda deney, DNA'daki bir nükleotid kırıldığında, proteinin her zaman, örtüşmeyen bir kod için tipik olan yalnızca bir amino asidi etkilediğini göstermiştir.
ГЦУГЦУГ ГЦУГЦУГ ГЦУГЦУГ
HCC HCC HCC UHC CUG HCC CUG UGC HCU CUG
*** *** *** *** *** ***
Alanin - Alanin Ala - Cys - Lei Ala - Lei - Lei - Ala - Lei
bir B C
örtüşmeyen kod örtüşen kod
Pirinç. 34. Genomda örtüşmeyen bir kodun varlığını açıklayan şema (metindeki açıklama).
Genetik kodun örtüşmemesi başka bir özellik ile ilişkilidir - bilginin okunması belirli bir noktadan başlar - başlatma sinyali. mRNA'daki böyle bir başlatma sinyali, AUG metionini kodlayan kodondur.
Bir kişinin hala genel kuraldan sapan ve örtüşen az sayıda gene sahip olduğuna dikkat edilmelidir.
e. Kompaktlık.
Kodonlar arasında noktalama işareti yoktur. Başka bir deyişle, üçüzler, örneğin anlamsız bir nükleotit tarafından birbirinden ayrılmaz. Genetik kodda "noktalama işaretlerinin" olmadığı deneylerle kanıtlanmıştır.
ve. çok yönlülük
Kod, Dünya'da yaşayan tüm organizmalar için aynıdır. Genetik kodun evrenselliğinin doğrudan kanıtı, DNA dizilerinin karşılık gelen protein dizileriyle karşılaştırılmasıyla elde edildi. Tüm bakteri ve ökaryotik genomlarda aynı kod değerlerinin kullanıldığı ortaya çıktı. İstisnalar var, ama çok değil.
Genetik kodun evrenselliğine ilişkin ilk istisnalar, bazı hayvan türlerinin mitokondrilerinde bulundu. Bu, amino asit triptofanı kodlayan UGG kodonu ile aynı şeyi okuyan sonlandırıcı kodon UGA ile ilgiliydi. Evrensellikten daha nadir başka sapmalar da bulunmuştur.
DNA kod sistemi.
DNA'nın genetik kodu 64 üçlü nükleotitten oluşur. Bu üçlülere kodon denir. Her kodon, protein sentezinde kullanılan 20 amino asitten birini kodlar. Bu, kodda bir miktar fazlalık sağlar: çoğu amino asit, birden fazla kodon tarafından kodlanır.
Bir kodon birbiriyle ilişkili iki işlevi yerine getirir: translasyonun başlangıcını işaret eder ve amino asit metioninin (Met) büyüyen polipeptit zincirine dahil edilmesini kodlar. DNA kod sistemi, genetik kodun RNA kodonları veya DNA kodonları olarak ifade edilebilmesi için tasarlanmıştır. RNA kodonları RNA'da (mRNA) meydana gelir ve bu kodonlar, polipeptitlerin sentezi sırasında (translasyon adı verilen bir süreç) bilgileri okuyabilir. Ancak her mRNA molekülü, karşılık gelen genden transkripsiyonda bir nükleotid dizisi alır.
İki amino asit (Met ve Trp) hariç tümü, 2 ila 6 farklı kodon tarafından kodlanabilir. Bununla birlikte, çoğu organizmanın genomu, belirli kodonların diğerlerine göre tercih edildiğini gösterir. Örneğin insanlarda alanin, GCC tarafından GCG'ye göre dört kat daha sık kodlanır. Bu muhtemelen bazı kodonlar için çeviri aygıtının (örneğin ribozom) daha yüksek bir çeviri etkinliğini gösterir.

Genetik kod neredeyse evrenseldir. Aynı kodonlar aynı amino asit dizisine atanır ve hayvanlarda, bitkilerde ve mikroorganizmalarda aynı başlama ve durma sinyalleri ezici bir çoğunlukla aynıdır. Ancak, bazı istisnalar bulunmuştur. Bunların çoğu, üç durdurma kodonundan bir veya ikisinin bir amino aside atanmasını içerir.
Rusya Federasyonu Eğitim ve Bilim Bakanlığı Federal Eğitim Ajansı
Yüksek Mesleki Eğitim Devlet Eğitim Kurumu "I.I. Polzunov'un adını taşıyan Altay Devlet Teknik Üniversitesi"
Doğa Bilimleri ve Sistem Analizi Bölümü
"Genetik kod" konulu makale
1. Genetik kod kavramı
3. Genetik bilgi
bibliyografya
1. Genetik kod kavramı
Genetik kod, nükleik asit moleküllerindeki kalıtsal bilgileri canlı organizmaların karakteristiği olan bir nükleotid dizisi şeklinde kaydetmek için tek bir sistemdir. Her nükleotid, parçası olan azotlu bazın adının başladığı bir büyük harfle gösterilir: - A (A) adenin; - G (G) guanin; - C(C) sitozin; - T (T) timin (DNA'da) veya U (U) urasil (mRNA'da).
Genetik kodun hücrede uygulanması iki aşamada gerçekleşir: transkripsiyon ve translasyon.
Bunlardan ilki çekirdekte gerçekleşir; DNA'nın karşılık gelen bölümlerinde mRNA moleküllerinin sentezinden oluşur. Bu durumda, DNA nükleotid dizisi, RNA nükleotid dizisine "yeniden yazılır". İkinci aşama sitoplazmada, ribozomlarda gerçekleşir; bu durumda, i-RNA'nın nükleotid dizisi, proteindeki amino asit dizisine çevrilir: bu aşama, transfer RNA'sının (t-RNA) ve karşılık gelen enzimlerin katılımıyla ilerler.
2. Genetik kodun özellikleri
1. Üçlülük
Her amino asit, 3 nükleotitlik bir dizi tarafından kodlanır.
Bir üçlü veya kodon, bir amino asidi kodlayan üç nükleotit dizisidir.
Kod monoplet olamaz, çünkü 4 (DNA'daki farklı nükleotidlerin sayısı) 20'den azdır. Kod çift olamaz, çünkü 16 (2 ile 4 nükleotidin kombinasyon ve permütasyon sayısı 20'den azdır. Kod üçlü olabilir, çünkü 64 (4'ten 3'e kadar olan kombinasyon ve permütasyon sayısı 20'den fazladır.)
2. Dejenerasyon.
Metiyonin ve triptofan dışındaki tüm amino asitler birden fazla üçlü tarafından kodlanır: 2 amino asit 1 üçlü = 2 9 amino asit her biri 2 üçlü = 18 1 amino asit 3 üçlü = 3 5 amino asit her biri 4 üçlü = 20 3 amino asit 6 üçlü her biri = 18 20 amino asit için toplam 61 üçlü kod.
3. İntergenik noktalama işaretlerinin varlığı.
Bir gen, bir polipeptit zincirini veya bir tRNA, rRNA veya sRNA molekülünü kodlayan bir DNA bölümüdür.
tRNA, rRNA ve sRNA genleri proteinleri kodlamaz.
Bir polipeptidi kodlayan her genin sonunda 3 sonlandırma kodonundan veya durdurma sinyalinden en az biri vardır: UAA, UAG, UGA. Yayını sonlandırıyorlar.
Geleneksel olarak, AUG kodonu ayrıca noktalama işaretlerine aittir - lider diziden sonraki ilk. Büyük harf işlevini yerine getirir. Bu pozisyonda formilmetiyonin (prokaryotlarda) kodlar.
4. Benzersizlik.
Her üçlü yalnızca bir amino asidi kodlar veya bir çeviri sonlandırıcıdır.
İstisna, AUG kodonudur. Prokaryotlarda, ilk konumda (büyük harf) formilmetiyonin için kodlar ve diğer herhangi bir konumda metionin için kodlar.
5. Kompaktlık veya intragenik noktalama işaretlerinin olmaması.
Bir gen içinde, her nükleotid, önemli bir kodonun parçasıdır.
1961'de Seymour Benzer ve Francis Crick deneysel olarak kodun üçlü ve kompakt olduğunu kanıtladı.

Deneyin özü: "+" mutasyon - bir nükleotidin eklenmesi. "-" mutasyon - bir nükleotidin kaybı. Bir genin başlangıcındaki tek bir "+" veya "-" mutasyonu tüm geni bozar. Bir çift "+" veya "-" mutasyon da tüm geni bozar. Genin başlangıcındaki üçlü bir "+" veya "-" mutasyon, genin sadece bir kısmını bozar. Dörtlü bir "+" veya "-" mutasyonu yine tüm geni bozar.
Deney, kodun üçlü olduğunu ve genin içinde noktalama işareti olmadığını kanıtlıyor. Deney, iki bitişik faj geni üzerinde gerçekleştirildi ve ek olarak, genler arasında noktalama işaretlerinin varlığını gösterdi.
3. Genetik bilgi
Genetik bilgi, bir organizmanın atalarından alınan ve kalıtsal yapılara genetik kod şeklinde gömülü olan özelliklerinin bir programıdır.
Genetik bilgi oluşumunun şemaya göre ilerlediği varsayılmaktadır: jeokimyasal süreçler - mineral oluşumu - evrimsel kataliz (otokataliz).
İlk ilkel genlerin mikrokristal kil kristalleri olması ve her yeni kil tabakasının, yapı hakkında bilgi alıyormuş gibi, bir öncekinin yapısal özelliklerine göre sıralanması mümkündür.
Genetik bilginin gerçekleştirilmesi, üç RNA'nın yardımıyla protein moleküllerinin sentezi sürecinde gerçekleşir: bilgi (mRNA), taşıma (tRNA) ve ribozomal (rRNA). Bilgi aktarımı süreci: - doğrudan iletişim kanalı aracılığıyla: DNA - RNA - protein; ve - geri besleme kanalı aracılığıyla: çevre - protein - DNA.
Canlı organizmalar bilgiyi alabilir, saklayabilir ve iletebilir. Ayrıca, canlı organizmalar kendileri ve çevrelerindeki dünya hakkında aldıkları bilgileri mümkün olduğunca verimli kullanma eğilimindedir. Genlere gömülü olan ve canlı bir organizmanın var olması, gelişmesi ve üremesi için gerekli olan kalıtsal bilgiler her bireyden sonraki nesillere aktarılır. Bu bilgi organizmanın gelişim yönünü belirler ve çevre ile etkileşimi sürecinde bireyine verilen tepki bozulabilir, böylece soyundan gelenlerin gelişiminin evrimini sağlar. Canlı bir organizmanın evrim sürecinde, bilginin değeri de dahil olmak üzere yeni bilgiler ortaya çıkar ve hatırlanır.
Kalıtsal bilgilerin belirli çevresel koşullar altında uygulanması sırasında, belirli bir biyolojik türün organizmalarının fenotipi oluşur.
Genetik bilgi vücudun morfolojik yapısını, büyümesini, gelişmesini, metabolizmasını, zihinsel deposunu, hastalıklara yatkınlığını ve genetik kusurlarını belirler.
Bilginin canlıların oluşumu ve evrimindeki rolünü haklı olarak vurgulayan birçok bilim adamı, bu durumu yaşamın ana kriterlerinden biri olarak kaydetti. Yani, V.I. Karagodin, "Canlı, bilginin ve onun kodladığı yapıların böyle bir varoluş biçimidir, bu bilginin uygun çevre koşullarında yeniden üretilmesini sağlar." Bilginin yaşamla bağlantısı da A.A. Lyapunov: "Yaşam, kalıcı reaksiyonlar geliştirmek için tek tek moleküllerin durumları tarafından kodlanan bilgileri kullanan, oldukça düzenli bir madde halidir." Tanınmış astrofizikçimiz N.S. Kardashev ayrıca yaşamın bilgi bileşenini de vurgulamaktadır: “Yaşam, çevre ve kendi yapıları hakkında en basit bilgileri, kendilerini korumak için kullandıkları ilk başta hatırlayabilen ve kullanabilen özel bir tür molekülü sentezleme olasılığı nedeniyle ortaya çıkar. , üreme için ve özellikle bizim için önemli olan daha fazla bilgi edinmek için." Ekolojist F. Tipler, "Ölümsüzlük Fiziği" adlı kitabında, canlı organizmaların bilgi depolama ve iletme yeteneğine dikkat çekiyor: "Hayatı, doğal seçilim tarafından korunan bir tür kodlanmış bilgi olarak tanımlıyorum." Ayrıca, eğer böyleyse, yaşam bilgi sisteminin ebedi, sonsuz ve ölümsüz olduğuna inanır.
Genetik kodun keşfi ve moleküler biyolojide kalıpların oluşturulması, modern genetiği ve Darwin'in evrim teorisini birleştirme ihtiyacını gösterdi. Böylece yeni bir biyolojik paradigma doğdu - zaten klasik olmayan biyoloji olarak kabul edilebilecek sentetik evrim teorisi (STE).
Darwin'in evriminin ana fikirleri - kalıtım, değişkenlik, doğal seleksiyon - yaşayan dünyanın evriminin modern görüşünde, sadece doğal seleksiyonla değil, aynı zamanda genetik olarak belirlenen bu tür seleksiyonla da desteklenir. Sentetik veya genel evrim gelişiminin başlangıcı, S.S.'nin eseri olarak kabul edilebilir. Chetverikov, bireysel özelliklerin ve bireylerin seçime tabi tutulmadığını, ancak tüm popülasyonun genotipinin, ancak bireysel bireylerin fenotipik özellikleri aracılığıyla gerçekleştirildiğinin gösterildiği popülasyon genetiği üzerine. Bu, faydalı değişikliklerin popülasyon boyunca yayılmasına yol açar. Böylece, evrim mekanizması hem genetik düzeyde rastgele mutasyonlar yoluyla hem de mutasyonel özelliklerin çevreye adaptasyonunu belirleyen en değerli özelliklerin (bilginin değeri!) .
Mevsimsel iklim değişiklikleri, çeşitli doğal veya insan kaynaklı afetler bir yandan popülasyonlarda gen tekrarlama sıklığının değişmesine ve bunun sonucunda kalıtsal değişkenliğin azalmasına neden olur. Bu sürece bazen genetik sürüklenme denir. Öte yandan, çeşitli mutasyonların konsantrasyonundaki değişikliklere ve popülasyonda bulunan genotiplerin çeşitliliğinde bir azalmaya, bu da seçim eyleminin yönünde ve yoğunluğunda değişikliklere yol açabilir.
4. İnsan genetik kodunun deşifre edilmesi
Mayıs 2006'da, insan genomunu deşifre etmek için çalışan bilim adamları, son eksik dizili insan kromozomu olan kromozom 1'in eksiksiz bir genetik haritasını yayınladılar.
İnsan Genom Projesinin resmi sonunu işaret eden bir ön insan genetik haritası 2003 yılında yayınlandı. Çerçevesi içinde, insan genlerinin %99'unu içeren genom fragmanları dizildi. Gen tanımlamasının doğruluğu %99,99'du. Ancak projenin sonunda 24 kromozomdan sadece dördü tam olarak dizilenebilmişti. Gerçek şu ki, kromozomlar genlere ek olarak, herhangi bir özelliği kodlamayan ve protein sentezinde yer almayan fragmanlar içerir. Bu parçaların organizmanın yaşamında oynadığı rol hala bilinmemektedir, ancak giderek daha fazla araştırmacı, çalışmalarının en yakın ilgiyi gerektirdiğine inanmaya meyillidir.
ders 5 Genetik Kod
kavram tanımı
Genetik kod, DNA'daki nükleotid dizisini kullanarak proteinlerdeki amino asit dizisi hakkında bilgi kaydetmek için bir sistemdir.
DNA, protein sentezinde doğrudan yer almadığından, kod RNA dilinde yazılmıştır. RNA, timin yerine urasil içerir.
Genetik kodun özellikleri
1. Üçlülük
Her amino asit, 3 nükleotitlik bir dizi tarafından kodlanır.
Tanım: Üçlü veya kodon, bir amino asidi kodlayan üç nükleotit dizisidir.
Kod monoplet olamaz, çünkü 4 (DNA'daki farklı nükleotidlerin sayısı) 20'den azdır. Kod çift olamaz, çünkü 16 (2 ile 4 nükleotidin kombinasyon ve permütasyon sayısı 20'den azdır. Kod üçlü olabilir, çünkü 64 (4'ten 3'e kadar olan kombinasyon ve permütasyon sayısı 20'den fazladır.)
2. Dejenerasyon.
Metionin ve triptofan hariç tüm amino asitler, birden fazla üçlü tarafından kodlanır:
1 üçlü için 2 AK = 2.
9 AK x 2 üçüz = 18.
1 AK 3 üçüz = 3.
5 AK x 4 üçüz = 20.
3 AK x 6 üçüz = 18.
20 amino asit için toplam 61 üçlü kod.
3. İntergenik noktalama işaretlerinin varlığı.
Tanım:
Gen bir polipeptit zincirini veya bir molekülü kodlayan bir DNA parçasıdır tPHK, rRNA veyasPHK.
genlertPHK, rPHK, sPHKproteinler kodlamaz.
Bir polipeptidi kodlayan her genin sonunda, RNA stop kodonlarını veya stop sinyallerini kodlayan 3 üçlüden en az biri bulunur. mRNA'da şöyle görünürler: UAA, UAG, UGA . Yayını sonlandırırlar (sonlandırırlar).
Geleneksel olarak, kodon noktalama işaretleri için de geçerlidir. Ağustos - lider dizisinden sonraki ilk. (Bkz. Ders 8) Büyük harf işlevini yerine getirir. Bu pozisyonda formilmetiyonin (prokaryotlarda) kodlar.
4. Benzersizlik.
Her üçlü yalnızca bir amino asidi kodlar veya bir çeviri sonlandırıcıdır.
Bunun istisnası kodondur. Ağustos . Prokaryotlarda, ilk konumda (büyük harf) formilmetiyonin için kodlar ve diğer herhangi bir konumda metionin için kodlar.
5. Kompaktlık veya intragenik noktalama işaretlerinin olmaması.
Bir gen içinde, her nükleotid, önemli bir kodonun parçasıdır.
1961'de Seymour Benzer ve Francis Crick deneysel olarak kodun üçlü ve kompakt olduğunu kanıtladılar.
Deneyin özü: "+" mutasyon - bir nükleotidin eklenmesi. "-" mutasyon - bir nükleotidin kaybı. Bir genin başlangıcındaki tek bir "+" veya "-" mutasyonu tüm geni bozar. Bir çift "+" veya "-" mutasyon da tüm geni bozar.
Genin başlangıcındaki üçlü bir "+" veya "-" mutasyon, genin sadece bir kısmını bozar. Dörtlü bir "+" veya "-" mutasyonu yine tüm geni bozar.
Deney bunu kanıtlıyor kod üçlüdür ve genin içinde noktalama işareti yoktur. Deney iki bitişik faj geni üzerinde gerçekleştirildi ve ayrıca şunu gösterdi: genler arasında noktalama işaretlerinin varlığı.
6. Çok yönlülük.
Genetik kod, Dünya'da yaşayan tüm canlılar için aynıdır.
1979'da Burrell açıldı ideal insan mitokondri kodu
Tanım:
"İdeal", yarı-çift kodun yozlaşma kuralının yerine getirildiği genetik koddur: İki üçlüdeki ilk iki nükleotit çakışırsa ve üçüncü nükleotidler aynı sınıfa aitse (her ikisi de pürin veya her ikisi de pirimidindir) , o zaman bu üçlüler aynı amino asidi kodlar .
Genel kodda bu kuralın iki istisnası vardır. Evrenseldeki ideal koddan her iki sapma da temel noktalarla ilgilidir: protein sentezinin başlangıcı ve sonu:
kodon | Evrensel kod | mitokondriyal kodlar |
|||
Omurgalılar | omurgasızlar | Maya | Bitkiler |
||
DUR | DUR |
||||
UA ile | |||||
bir G A | DUR | ||||
DUR | 230 ikame, kodlanan amino asidin sınıfını değiştirmez. yırtılabilirliğe. 1956'da Georgy Gamov, örtüşen kodun bir varyantını önerdi. Gamow koduna göre, gendeki üçüncüden başlayarak her nükleotit, 3 kodonun bir parçasıdır. Genetik kod deşifre edildiğinde, örtüşmediği ortaya çıktı, yani. her nükleotid sadece bir kodonun parçasıdır. Örtüşen genetik kodun avantajları: kompaktlık, protein yapısının bir nükleotidin eklenmesine veya silinmesine daha az bağımlı olması. Dezavantaj: Protein yapısının nükleotid ikamesine yüksek bağımlılığı ve komşulara kısıtlama. 1976'da φX174 fajının DNA'sı dizilendi. 5375 nükleotitten oluşan tek sarmallı dairesel bir DNA'ya sahiptir. Fajın 9 proteini kodladığı biliniyordu. Bunlardan 6 tanesinde birbiri ardına yer alan genler tespit edildi. Bir örtüşme olduğu ortaya çıktı. E geni tamamen genin içindedir. D . Başlangıç kodonu, okumadaki bir nükleotid kaymasının bir sonucu olarak ortaya çıkar. Gen J genin bittiği yerde başlar D . Gen başlatma kodonu J genin sonlandırma kodonu ile örtüşür. D iki nükleotidin yer değiştirmesinden kaynaklanır. Tasarım, üçün katı olmayan bir dizi nükleotid tarafından "okuma çerçevesi kayması" olarak adlandırılır. Bugüne kadar, örtüşme yalnızca birkaç faj için gösterilmiştir. DNA'nın bilgi kapasitesi Dünyada 6 milyar insan var. Onlar hakkında kalıtsal bilgiler 4x10 13 kitap sayfası. Bu sayfalar 6 NSU binasının yerini alacaktı. 6x109 sperm bir yüksüğün yarısını alır. DNA'ları yüksüğün dörtte birinden daha az yer kaplar. | ||||
Protein sentezlemek gerekirse, hücrenin önünde ciddi bir sorun ortaya çıkar - DNA'daki bilgiler kodlanmış bir dizi şeklinde depolanır. 4 karakter(nükleotidler) ve proteinlerden oluşur. 20 farklı karakter(amino asitler). Amino asitleri kodlamak için dört sembolü aynı anda kullanmaya çalışırsanız, 20 proteinojenik amino asit varken sadece 16 kombinasyon elde edersiniz.Yeterli değil ...
Bu konuda dahiyane bir düşünce örneği var:
"Örneğin, sadece kartın rengine dikkat ettiğimiz bir iskambil destesini ele alalım. Aynı türden kaç tane üçüz elde edilebilir? Elbette dört: üç kupa, üç karo, üç maça ve üç sopa. Aynı takımdan ve biri diğerinden iki kartla kaç tane üçüz vardır? Diyelim ki üçüncü kart için dört seçeneğimiz var. Yani 4x3 = 12 olasılığımız var. Ek olarak, üç farklı karta sahip dört üçlümüz var. Yani, 4 + 12 + 4 = 20 ve bu, almak istediğimiz tam amino asit sayısıdır "(George Gamow, İngiliz George Gamow, 1904-1968, Sovyet ve Amerikan teorik fizikçi, astrofizikçi ve bilimin popülerleştiricisi).
Gerçekten de deneyler, her amino asit için iki zorunlu nükleotid ve daha az spesifik olan üçüncü bir değişken (" sallanma etkisi"). Dört karakterden üçünü alırsanız, amino asitlerin sayısıyla büyük ölçüde örtüşen 64 kombinasyon elde edersiniz. Böylece, herhangi bir amino asidin üç nükleotit tarafından kodlandığı bulundu. Bu üçlüye denir. kodon. Daha önce de belirtildiği gibi, 64 seçenek var. Üçü herhangi bir amino asidi kodlamaz, bunlara "sözde" denir. anlamsız kodonlar"(Fransızca. anlamsız- saçmalık) veya "kodonları durdur".
Genetik Kod
Genetik (biyolojik) bir kod, proteinlerin yapısı hakkındaki bilgileri bir nükleotid dizisi biçiminde kodlamanın bir yoludur. Dört basamaklı nükleotit dilini (A, G, U, C) yirmi basamaklı bir amino asit diline çevirmek için tasarlanmıştır. Karakteristik özelliklere sahiptir:
- üçlülükÜç nükleotid, bir amino asidi kodlayan bir kodon oluşturur. Toplamda 61 duyu kodonu vardır.
- özgüllük(veya benzersizlik) - her kodon sadece bir amino aside karşılık gelir.
- yozlaşma Bir amino asit birden fazla kodona sahip olabilir.
- çok yönlülük- biyolojik kod dünyadaki tüm organizma türleri için aynıdır (ancak memelilerin mitokondrilerinde istisnalar vardır).
- eşdoğrusallık- kodonların dizisi, kodlanmış proteindeki amino asitlerin dizisine karşılık gelir.
- örtüşmeyen- yan yana yerleştirilmiş üçüzler birbiri üzerine binmez.
- Eksik noktalama– üçüzler arasında ek nükleotid veya başka sinyal yoktur.
- Tek yönlü- protein sentezi sırasında, kodon okuması, boşluklar veya geri izleme olmaksızın sırayla ilerler.
Bununla birlikte, biyolojik kodun, bir geçiş işlevi gören ek moleküller olmadan kendini gösteremeyeceği açıktır. adaptör işlevi.
Transfer RNA'larının adaptör rolü
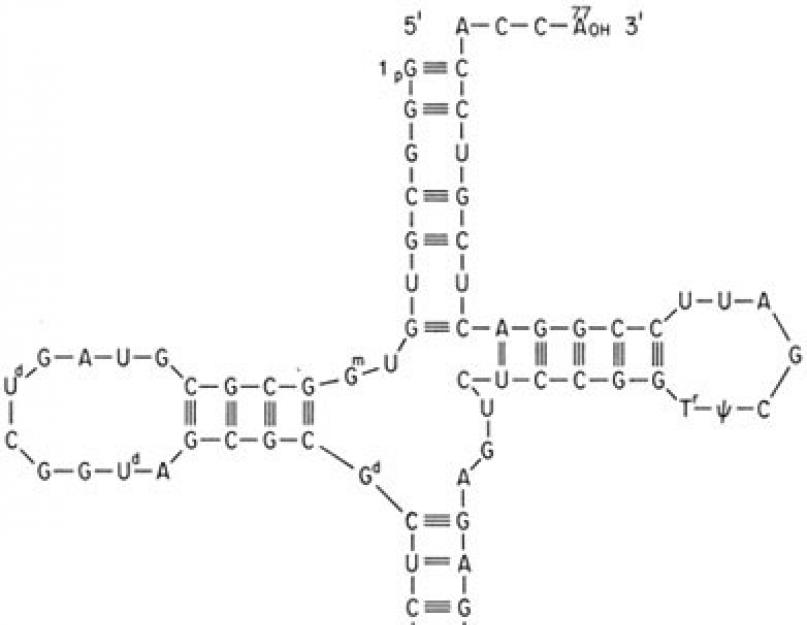
Transfer RNA'ları, 4 harfli bir nükleik asit dizisi ile 20 harfli bir protein dizisi arasındaki tek aracıdır.
Her transfer RNA'sı, antikodon döngüsünde belirli bir üçlü diziye sahiptir ( antikodon) ve yalnızca o antikodona uyan bir amino asit ekleyebilir. Protein molekülüne hangi amino asidin dahil edileceğini belirleyen tRNA'da bir veya başka bir antikodonun varlığıdır, tk. ne ribozom ne de mRNA amino asidi tanımaz.
Böylece, tRNA'nın bağdaştırıcı rolü dır-dir:
- amino asitlere spesifik bağlanmada,
- spesifik olarak, kodon-antikodon etkileşimine göre, mRNA'ya bağlanma,
- ve sonuç olarak, mRNA bilgisine uygun olarak amino asitlerin protein zincirine dahil edilmesinde.
Bir amino asidin tRNA'ya bağlanması bir enzim tarafından gerçekleştirilir. aminoasil-tRNA sentetaz, aynı anda iki bileşik için özgüllüğe sahiptir: herhangi bir amino asit ve buna karşılık gelen tRNA. Reaksiyon, iki makroerjik ATP bağı gerektirir. Amino asit, a-karboksil grubu aracılığıyla tRNA alıcı döngüsünün 3' ucuna bağlanır ve amino asit ile tRNA arasındaki bağ, makroerjik. α-amino grubu serbest kalır.
Aminoasil-tRNA sentez reaksiyonu
Yaklaşık 60 farklı tRNA olduğu için, bazı amino asitler iki veya daha fazla tRNA'ya karşılık gelir. Aynı amino asidi bağlayan farklı tRNA'lara denir. izoalıcı.
Vücudun metabolizmasında başrol
proteinlere ve nükleik asitlere aittir.
Protein maddeleri, tüm hayati hücre yapılarının temelini oluşturur, alışılmadık derecede yüksek reaktiviteye sahiptir ve katalitik fonksiyonlara sahiptir.
Nükleik asitler hücrenin en önemli organının bir parçasıdır - çekirdeğin yanı sıra sitoplazma, ribozomlar, mitokondri vb. Nükleik asitler kalıtım, vücut değişkenliği ve protein sentezinde önemli ve birincil bir rol oynar.
Plan sentez protein, hücre çekirdeğinde depolanır ve doğrudan sentez, çekirdeğin dışında gerçekleşir, bu nedenle gereklidir. teslimat hizmeti kodlanmış plan çekirdekten sentez yerine. Bu dağıtım hizmeti RNA molekülleri tarafından gerçekleştirilir.
süreç başlar çekirdek hücreler: DNA "merdiveninin" bir kısmı gevşer ve açılır. Bu nedenle, RNA harfleri, DNA zincirlerinden birinin açık DNA harfleriyle bağlar oluşturur. Enzim, onları bir ipliğe bağlamak için RNA'nın harflerini aktarır. Böylece DNA'nın harfleri, RNA'nın harflerine "yeniden yazılır". Yeni oluşan RNA zinciri ayrılır ve DNA "merdiveni" tekrar kıvrılır. DNA'dan bilgi okuma ve RNA şablonunu sentezleme işlemine denir. transkripsiyon ve sentezlenen RNA bilgisel veya i-RNA .
Daha fazla modifikasyondan sonra, bu tür kodlanmış mRNA hazırdır. i-RNA çekirdekten çıkar ve i-RNA harflerinin deşifre edildiği protein sentezi bölgesine gider. i-RNA'nın üç harfinden oluşan her bir set, belirli bir amino asidi temsil eden bir "harf" oluşturur.
Başka bir RNA türü ise bu amino asidi arar, bir enzim yardımıyla yakalar ve protein sentezi bölgesine iletir. Bu RNA'ya transfer RNA veya tRNA denir. mRNA mesajı okunup çevrildikçe amino asit zinciri büyür. Bu zincir kıvrılarak benzersiz bir şekle bürünerek bir tür protein oluşturur. Protein katlama süreci bile dikkate değerdir: tüm hesaplamaları bir bilgisayarla yapmak. seçenekler 100 amino asitten oluşan orta boy bir proteinin katlanması 1027 (!) yıl alacaktır. Ve vücutta 20 amino asitlik bir zincirin oluşması bir saniyeden fazla sürmez ve bu süreç vücudun tüm hücrelerinde sürekli olarak gerçekleşir.
Genler, genetik kod ve özellikleri.
Dünya'da yaklaşık 7 milyar insan yaşıyor. 25-30 milyon tek yumurta ikizi dışında genetik olarak tüm insanlar farklıdır : her biri benzersizdir, benzersiz kalıtsal özelliklere, karakter özelliklerine, yeteneklerine, mizacına sahiptir.
Bu tür farklılıklar açıklanmıştır genotiplerdeki farklılıklar- bir organizmanın gen setleri; her biri benzersizdir. Belirli bir organizmanın genetik özellikleri somutlaştırılmıştır. proteinlerde - sonuç olarak, bir kişinin proteininin yapısı, biraz da olsa, başka bir kişinin proteininden farklıdır.
Bu demek değil insanların tam olarak aynı proteinlere sahip olmadığıdır. Aynı işlevleri yerine getiren proteinler aynı olabilir veya birbirinden bir veya iki amino asit bakımından çok az farklı olabilir. Fakat bulunmuyor tüm proteinlerin olacağı insanların Dünyasında (tek yumurta ikizleri hariç), aynıdır .
Bir proteinin birincil yapısı hakkında bilgi bir DNA molekülünün bir bölümünde bir dizi nükleotit olarak kodlanmış, gen - bir organizmanın kalıtsal bilgi birimi. Her DNA molekülü birçok gen içerir. Bir organizmanın tüm genlerinin toplamı, onun yapısını oluşturur. genotip . Böylece,
Gen, bir organizmanın DNA'nın ayrı bir bölümüne karşılık gelen kalıtsal bilgi birimidir.
Kalıtsal bilgiler kullanılarak kodlanır genetik Kod tüm organizmalar için evrensel olan ve yalnızca belirli organizmaların proteinlerini kodlayan ve genleri oluşturan nükleotitlerin değişiminde farklılık gösteren .
Genetik Kod farklı dizilerde (AAT, HCA, ACG, THC, vb.) birleştirilmiş DNA nükleotitlerinin üçlülerinden (üçlülerinden) oluşur ve bunların her biri belirli bir amino asidi (polipeptit zincirinde oluşturulacak) kodlar.
Aslında kod
sayar bir i-RNA molekülündeki nükleotid dizisi
, çünkü DNA'dan bilgiyi kaldırır (süreç transkripsiyonlar
) ve sentezlenmiş proteinlerin moleküllerindeki bir amino asit dizisine çevirir (işlem yayınlar
).
MRNA'nın bileşimi, üçlüleri olarak adlandırılan A-C-G-U nükleotitlerini içerir. kodonlar
: mRNA üzerindeki CHT DNA üçlüsü HCA üçlüsü olacak ve AAG DNA üçlüsü UUC üçlüsü olacak. Aynen öyle i-RNA kodonları
kayıttaki genetik kodu yansıtır.
Böylece, genetik kod - nükleik asit moleküllerindeki kalıtsal bilgileri bir nükleotid dizisi şeklinde kaydetmek için birleşik bir sistem . Genetik kod, azotlu bazlarda farklılık gösteren sadece dört nükleotid harfinden oluşan bir alfabenin kullanımına dayanmaktadır: A, T, G, C.
Genetik kodun ana özellikleri:

1. Genetik Kod üçlü. Üçlü (kodon), bir amino asidi kodlayan üç nükleotit dizisidir. Proteinler 20 amino asit içerdiğinden, her birinin bir nükleotid tarafından kodlanamayacağı açıktır ( DNA'da sadece dört tip nükleotit bulunduğundan, bu durumda 16 amino asit kodlanmamış kalır.). Amino asitleri kodlamak için iki nükleotid de yeterli değildir, çünkü bu durumda sadece 16 amino asit kodlanabilir. Bu, bir amino asidi kodlayan en küçük nükleotid sayısının en az üç olması gerektiği anlamına gelir. Bu durumda olası nükleotid üçlülerinin sayısı 43 = 64'tür.
2. Artıklık (yozlaşma) Kod, üçlü yapısının bir sonucudur ve bir amino asidin, yalnızca bir amino asit tarafından kodlanan metiyonin ve triptofan hariç, birkaç üçlü tarafından (20 amino asit olduğundan ve 64 üçlü olduğundan) kodlanabileceği anlamına gelir. üçlü. Ek olarak, bazı üçlüler belirli işlevleri yerine getirir: mRNA molekülünde UAA, UAG, UGA üçlüleri sonlandırıcı kodonlardır, yani. Dur-polipeptit zincirinin sentezini durduran sinyaller. DNA zincirinin başında bulunan metiyonine (AUG) karşılık gelen üçlü, bir amino asidi kodlamaz, ancak okuma (uyarıcı) başlatma işlevini yerine getirir.
3. belirsizlik kod - artıklık ile birlikte, kodun özelliği vardır benzersizlik : her kodon yalnızca eşleşir bir spesifik amino asit.
4. eşdoğrusallık kod, yani bir gendeki nükleotid dizisi kesinlikle proteindeki amino asitlerin sırasına karşılık gelir.
5. Genetik kod örtüşmeyen ve kompakt , yani "noktalama işaretleri" içermez. Bu, okuma işleminin, üst üste binen sütunların (üçlüler) olasılığına izin vermediği ve belirli bir kodondan başlayarak, okumanın sürekli olarak üçe üçe gittiği anlamına gelir. Dur-sinyaller ( sonlandırma kodonları).
6. Genetik kod evrensel yani, tüm organizmaların nükleer genleri, bu organizmaların organizasyon düzeyi ve sistematik konumu ne olursa olsun, proteinler hakkındaki bilgileri aynı şekilde kodlar.
Mevcut genetik kod tabloları şifre çözme için kodonlar i-RNA ve protein moleküllerinin yapı zincirleri.

Matris sentez reaksiyonları.
Canlı sistemlerde cansız doğada bilinmeyen reaksiyonlar vardır - matris sentez reaksiyonları.
"Matriks" terimi teknolojide madeni para, madalya, tipografik tip dökümü için kullanılan formu belirtirler: sertleştirilmiş metal, döküm için kullanılan formun tüm ayrıntılarını tam olarak yeniden üretir. matris sentezi bir matris üzerindeki döküme benzer: yeni moleküller, halihazırda var olan moleküllerin yapısında ortaya konan plana tam olarak uygun olarak sentezlenir.
Matris ilkesi yatıyor merkezde nükleik asitlerin ve proteinlerin sentezi gibi hücrenin en önemli sentetik reaksiyonları. Bu reaksiyonlarda, sentezlenen polimerlerdeki monomerik birimlerin kesin, kesin olarak spesifik bir dizisi sağlanır.
Burası yönlü monomerleri belirli bir yere çekmek hücreler - reaksiyonun gerçekleştiği bir matris görevi gören moleküllere dönüşür. Bu tür reaksiyonlar, moleküllerin rastgele çarpışması sonucu meydana gelseydi, sonsuz yavaş ilerlerlerdi. Matriks prensibine dayalı karmaşık moleküllerin sentezi hızlı ve doğru bir şekilde gerçekleştirilir. matrisin rolü matriks reaksiyonlarında oynayan nükleik asitlerin makromolekülleri DNA veya RNA .

monomerik moleküller, polimerin sentezlendiği - nükleotitler veya amino asitler - tamamlayıcılık ilkesine göre, matris üzerinde kesin olarak tanımlanmış, önceden belirlenmiş bir sırayla düzenlenir ve sabitlenir.
Sonra gelir monomer birimlerinin bir polimer zincirine "çapraz bağlanması" ve bitmiş polimer matristen düşürülür.
bundan sonra matris hazır yeni bir polimer molekülünün montajına. Belirli bir kalıba yalnızca bir madeni para, bir harf dökülebileceği gibi, belirli bir matris molekülü üzerinde yalnızca bir polimerin "birleştirilebileceği" açıktır.
Matris tipi reaksiyon- canlı sistemlerin kimyasının belirli bir özelliği. Tüm canlıların temel özelliğinin temelidir - kendi türünü yeniden üretme yeteneği.
Matris sentez reaksiyonları
1. DNA kopyalama - replikasyon (lat. replikasyondan - yenilemeden) - ana DNA molekülünün matrisi üzerinde bir kız deoksiribonükleik asit molekülünün sentez süreci. Ana hücrenin sonraki bölünmesi sırasında, her yavru hücre, orijinal ana hücrenin DNA'sı ile aynı olan bir DNA molekülünün bir kopyasını alır. Bu süreç, genetik bilginin nesilden nesile doğru bir şekilde iletilmesini sağlar. DNA replikasyonu, adı verilen 15-20 farklı proteinden oluşan karmaşık bir enzim kompleksi tarafından gerçekleştirilir. replisome . Sentez malzemesi, hücrelerin sitoplazmasında bulunan serbest nükleotidlerdir. Replikasyonun biyolojik anlamı, normalde somatik hücrelerin bölünmesi sırasında meydana gelen kalıtsal bilginin ana molekülden yavru moleküllere tam olarak aktarılmasında yatmaktadır.
DNA molekülü birbirini tamamlayan iki zincirden oluşur. Bu zincirler, enzimler tarafından kırılabilen zayıf hidrojen bağları ile bir arada tutulur. DNA molekülü kendini ikiye katlama (kopyalama) yeteneğine sahiptir ve molekülün her eski yarısında yeni bir yarısı sentezlenir.
Ek olarak, bir DNA molekülü üzerinde bir mRNA molekülü sentezlenebilir, bu daha sonra DNA'dan alınan bilgiyi protein sentezi bölgesine aktarır.
Bilgi aktarımı ve protein sentezi, bir matbaadaki matbaanın çalışmasına benzer bir matris ilkesini takip eder. DNA'dan gelen bilgiler tekrar tekrar kopyalanır. Kopyalama sırasında hatalar meydana gelirse, sonraki tüm kopyalarda tekrarlanacaktır.

Doğru, bir DNA molekülü tarafından bilgi kopyalamadaki bazı hatalar düzeltilebilir - hataları ortadan kaldırma işlemine denir tazminat. Bilgi aktarımı sürecindeki tepkimelerden ilki DNA molekülünün replikasyonu ve yeni DNA ipliklerinin sentezidir.
2. Transkripsiyon (Latince transkript - yeniden yazmadan) - tüm canlı hücrelerde meydana gelen, DNA'yı bir şablon olarak kullanarak RNA sentezi süreci. Başka bir deyişle, genetik bilginin DNA'dan RNA'ya aktarılmasıdır.
Transkripsiyon, DNA'ya bağımlı RNA polimeraz enzimi tarafından katalize edilir. RNA polimeraz, DNA molekülü boyunca 3 " → 5" yönünde hareket eder. Transkripsiyon adımlardan oluşur başlatma, uzama ve sonlandırma . Transkripsiyon birimi, aşağıdakilerden oluşan DNA molekülünün bir parçası olan operondur. promotör, kopyalanan kısım ve terminatör . i-RNA bir zincirden oluşur ve i-RNA molekülünün sentezinin başlangıcını ve sonunu aktive eden bir enzimin katılımıyla tamamlayıcılık kuralına uygun olarak DNA üzerinde sentezlenir.
Bitmiş mRNA molekülü, polipeptit zincirlerinin sentezinin gerçekleştiği ribozomlardaki sitoplazmaya girer.
3. Yayın (lat'den. tercüme- transfer, hareket) - ribozom tarafından gerçekleştirilen bilgi (matris) RNA (mRNA, mRNA) matrisindeki amino asitlerden protein sentezi süreci. Başka bir deyişle, i-RNA'nın nükleotid dizisinde yer alan bilgilerin polipeptiddeki amino asit dizisine çevrilmesi işlemidir.

4. ters transkripsiyon tek sarmallı RNA'dan gelen bilgilere dayalı olarak çift sarmallı DNA oluşturma işlemidir. Bu işleme ters transkripsiyon denir, çünkü genetik bilgi aktarımı, transkripsiyona göre “ters” yönde gerçekleşir. Ters transkripsiyon fikri, DNA'nın RNA'ya kopyalandığını ve daha sonra proteinlere çevrildiğini varsayan moleküler biyolojinin merkezi dogmasına karşı çıktığı için başlangıçta çok popüler değildi.
 Ancak 1970 yılında Temin ve Baltimore birbirinden bağımsız olarak ters transkriptaz (revertaz)
ve ters transkripsiyon olasılığı nihayet doğrulandı. 1975'te Temin ve Baltimore, Nobel Fizyoloji veya Tıp Ödülü'ne layık görüldü. Bazı virüsler (HIV enfeksiyonuna neden olan insan immün yetmezlik virüsü gibi), RNA'yı DNA'ya kopyalama yeteneğine sahiptir. HIV, DNA'ya entegre olan bir RNA genomuna sahiptir. Sonuç olarak, virüsün DNA'sı, konakçı hücrenin genomu ile birleştirilebilir. RNA'dan DNA sentezinden sorumlu ana enzime denir. tersine çevirmek. Tersazın işlevlerinden biri oluşturmaktır. tamamlayıcı DNA
(cDNA) viral genomdan. İlişkili enzim ribonükleaz, RNA'yı parçalar ve reverstase, DNA çift sarmalından cDNA'yı sentezler. cDNA, integraz yoluyla konak hücre genomuna entegre edilir. sonuç konak hücre tarafından viral proteinlerin sentezi yeni virüsler oluşturur. HIV durumunda, T-lenfositlerin apoptozu (hücre ölümü) de programlanır. Diğer durumlarda, hücre bir virüs dağıtıcısı olarak kalabilir.
Ancak 1970 yılında Temin ve Baltimore birbirinden bağımsız olarak ters transkriptaz (revertaz)
ve ters transkripsiyon olasılığı nihayet doğrulandı. 1975'te Temin ve Baltimore, Nobel Fizyoloji veya Tıp Ödülü'ne layık görüldü. Bazı virüsler (HIV enfeksiyonuna neden olan insan immün yetmezlik virüsü gibi), RNA'yı DNA'ya kopyalama yeteneğine sahiptir. HIV, DNA'ya entegre olan bir RNA genomuna sahiptir. Sonuç olarak, virüsün DNA'sı, konakçı hücrenin genomu ile birleştirilebilir. RNA'dan DNA sentezinden sorumlu ana enzime denir. tersine çevirmek. Tersazın işlevlerinden biri oluşturmaktır. tamamlayıcı DNA
(cDNA) viral genomdan. İlişkili enzim ribonükleaz, RNA'yı parçalar ve reverstase, DNA çift sarmalından cDNA'yı sentezler. cDNA, integraz yoluyla konak hücre genomuna entegre edilir. sonuç konak hücre tarafından viral proteinlerin sentezi yeni virüsler oluşturur. HIV durumunda, T-lenfositlerin apoptozu (hücre ölümü) de programlanır. Diğer durumlarda, hücre bir virüs dağıtıcısı olarak kalabilir.
Protein biyosentezindeki matris reaksiyonlarının sırası bir diyagram olarak gösterilebilir.

Böylece, protein biyosentezi- bu, DNA genlerinde kodlanan kalıtsal bilgilerin protein moleküllerinde belirli bir amino asit dizisinde gerçekleştirildiği plastik değişim türlerinden biridir.
Protein molekülleri esas olarak polipeptit zincirleri bireysel amino asitlerden oluşur. Ancak amino asitler kendi başlarına birbirleriyle bağlantı kuracak kadar aktif değillerdir. Bu nedenle amino asitler birbirleriyle birleşip bir protein molekülü oluşturmadan önce etkinleştirmek . Bu aktivasyon, özel enzimlerin etkisi altında gerçekleşir.
Aktivasyonun bir sonucu olarak, amino asit daha kararsız hale gelir ve aynı enzimin etkisi altında t-'ye bağlanır. RNA. Her amino asit kesinlikle spesifik bir t- RNA"kendi" amino asidini bulan ve dayanır ribozoma girer.
Bu nedenle, ribozom çeşitli alır aktifleştirilmiş amino asitler t- RNA. ribozom şuna benzer konveyör içine giren çeşitli amino asitlerden bir protein zinciri oluşturmak.
Kendi amino asidinin "oturduğu" t-RNA ile eşzamanlı olarak, " sinyal» çekirdekte bulunan DNA'dan. Bu sinyale göre, ribozomda bir veya başka bir protein sentezlenir.
DNA'nın protein sentezi üzerindeki yönlendirici etkisi doğrudan değil, özel bir aracı yardımıyla gerçekleştirilir - matris veya haberci RNA (mRNA veya i-RNA), Hangi çekirdeğe sentezlenir DNA'dan etkilenmez, bu nedenle bileşimi DNA'nın bileşimini yansıtır. RNA molekülü, adeta DNA formundan bir dökümdür. Sentezlenen mRNA ribozoma girer ve olduğu gibi onu bu yapıya aktarır. plan- Belirli bir proteini sentezlemek için ribozoma giren aktive amino asitlerin birbirleriyle hangi sırayla birleştirilmesi gerekir. Aksi halde, DNA'da kodlanan genetik bilgi önce mRNA'ya sonra da proteine aktarılır..
mRNA molekülü ribozoma girer ve yanıp söner o. Şu anda ribozomda bulunan bölümü belirlenir. kodon (üçlü), kendisine uygun bir yapı ile tamamen spesifik bir şekilde etkileşime girer. üçlü (antikodon) amino asidi ribozoma getiren transfer RNA'sında.
Amino asidi ile transfer RNA, belirli bir mRNA kodonuna yaklaşır ve bağlanır onunla; i-RNA'nın bir sonraki komşu sitesine başka bir tRNA'yı farklı bir amino asitle birleştirir ve böylece tüm i-RNA zinciri okunana kadar, tüm amino asitler uygun sırayla dizilerek bir protein molekülü oluşturana kadar. Ve amino asidi polipeptit zincirinin belirli bir bölgesine ileten t-RNA, amino asidinden arındırılmış ve ribozomdan çıkar.
Sonra tekrar sitoplazmada istenen amino asit ona katılabilir ve tekrar ribozoma aktarır. Protein sentezi sürecinde, bir değil, birkaç ribozom, poliribozom aynı anda dahil olur.
Genetik bilgi transferinin ana aşamaları:
1. Bir mRNA şablonunda olduğu gibi DNA üzerinde sentez (transkripsiyon)
2. i-RNA'nın içerdiği programa göre ribozomlardaki polipeptit zincirinin sentezi (çeviri)
.
Aşamalar tüm canlılar için evrenseldir, ancak bu süreçlerin zamansal ve mekansal ilişkileri pro- ve ökaryotlarda farklılık gösterir.
saat prokaryotlar DNA sitoplazmada bulunduğundan transkripsiyon ve translasyon aynı anda gerçekleşebilir. saat ökaryot transkripsiyon ve translasyon, uzay ve zamanda kesin olarak ayrılır: çekirdekte çeşitli RNA'ların sentezi meydana gelir, bundan sonra RNA moleküllerinin nükleer zardan geçerek çekirdeği terk etmesi gerekir. RNA daha sonra sitoplazmada protein sentezi bölgesine taşınır.



