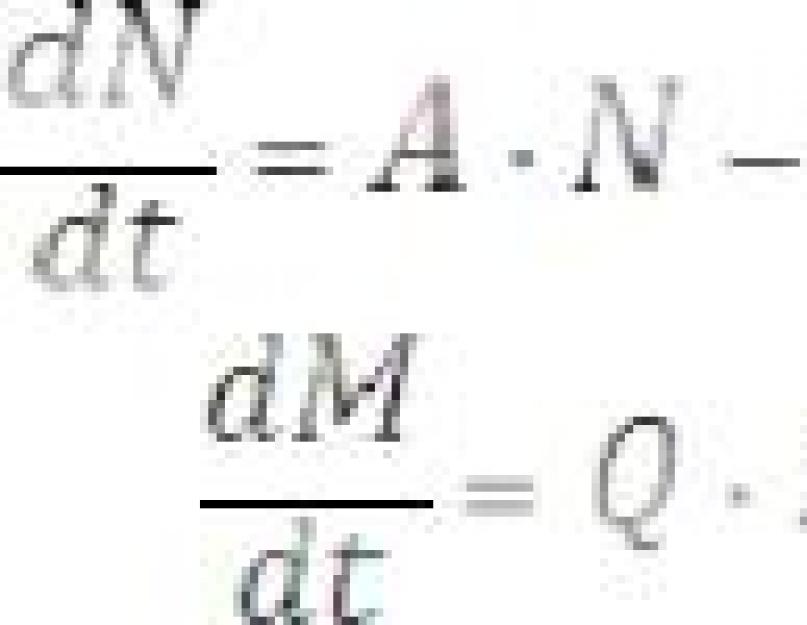
Federal ajans eğitimin
Durum Eğitim kurumu
daha yüksek mesleki Eğitim
"İjevsk Devleti Teknik Üniversite»
Uygulamalı Matematik Fakültesi
“Süreç ve teknolojilerin matematiksel modellenmesi” Bölümü
"Diferansiyel Denklemler" disiplininde
Konu: “Yırtıcı-av modelinin nitel araştırması”
İjevsk 2010
GİRİİŞ
1. “YIRICI-KURBAN” MODELİNİN PARAMETRELERİ VE TEMEL DENKLEMLERİ
2.2 “Yırtıcı-av” tipinin genelleştirilmiş Voltaire modelleri.
3. “YIRTICI-KURBAN” MODELİNİN PRATİK UYGULAMASI
ÇÖZÜM
KAYNAKÇA
GİRİİŞ
Günümüzde çevre sorunları büyük önem taşımaktadır. Önemli bir adım Bu sorunların çözümü ekolojik sistemlerin matematiksel modellerinin geliştirilmesidir.
Ekolojinin temel görevlerinden biri modern sahne doğal sistemlerin yapısı ve işleyişinin incelenmesi, araştırılmasıdır. genel desenler. Matematiğin ekoloji üzerinde büyük etkisi oldu ve özellikle diferansiyel denklemler teorisi, kararlılık teorisi ve optimal kontrol teorisi gibi bölümler matematiksel ekolojinin oluşumuna katkıda bulundu.
Matematiksel ekoloji alanındaki ilk çalışmalardan biri A.D. Lotki (1880 - 1949), yırtıcı-av ilişkileriyle birbirine bağlanan farklı popülasyonların etkileşimini tanımlayan ilk kişiydi. Yırtıcı-av modelinin çalışmasına büyük katkı V. Volterra (1860 - 1940), V.A. Kostitsyn (1883-1963) Günümüzde popülasyonların etkileşimini açıklayan denklemlere Lotka-Volterra denklemleri adı verilmektedir.
Lotka-Volterra denklemleri ortalama değerlerin (nüfus büyüklüğü) dinamiklerini tanımlar. Şu anda, bunlara dayanarak, integral diferansiyel denklemlerle tanımlanan daha genel popülasyon etkileşimi modelleri oluşturulmuş ve kontrollü avcı-av modelleri üzerinde çalışılmaktadır.
Matematiksel ekolojinin önemli sorunlarından biri ekosistemlerin istikrarı ve bu sistemlerin yönetimi sorunudur. Yönetim, bir sistemi kullanım veya restorasyon amacıyla bir kararlı durumdan diğerine aktarmak amacıyla gerçekleştirilebilir.
1. PREDATOR-PRİMAT MODELİNİN PARAMETRELERİ VE TEMEL DENKLEMLERİ
Denemeler matematiksel modelleme etkileşim halindeki popülasyonlar da dahil olmak üzere hem bireysel biyolojik popülasyonların hem de toplulukların dinamikleri çeşitli türler uzun zamandan beri yürütülmektedir. İzole nüfus artışının ilk modellerinden biri (2.1), 1798'de Thomas Malthus tarafından önerildi:
, (1.1)Bu model aşağıdaki parametrelerle belirtilir:
N - popülasyon büyüklüğü;
- Doğum ve ölüm oranları arasındaki fark.Bu denklemi entegre edersek şunu elde ederiz:
, (1.2)burada N(0), t = 0 anındaki popülasyon büyüklüğüdür. Açıkçası, Malthus modeli
> 0, bu büyümeyi sağlayan kaynakların her zaman sınırlı olduğu doğal popülasyonlarda asla gözlemlenmeyen sonsuz bir sayı artışı sağlar. Flora ve fauna popülasyonlarının sayısındaki değişiklikler açıklanamaz basit yasa Malthus'a göre, büyüme dinamikleri birbiriyle ilişkili birçok nedenden etkilenir - özellikle her türün üremesi, bu türün evrim sürecinde korunmasını sağlayacak şekilde kendi kendini düzenler ve değiştirilir.Bu modellerin matematiksel tanımları, bitki ve hayvan organizmalarının kendi aralarında ve çevreyle oluşturdukları toplulukların ilişkilerinin bilimi olan matematiksel ekoloji tarafından ele alınmaktadır.
Farklı türlerin çeşitli popülasyonlarını içeren biyolojik topluluk modellerine ilişkin en ciddi çalışma İtalyan matematikçi Vito Volterra tarafından gerçekleştirildi:
, - popülasyon boyutu; - oranlar doğal artış nüfusun (veya ölüm oranının); - türler arası etkileşim katsayıları. Katsayıların seçimine bağlı olarak model, ya türlerin ortak bir kaynak için mücadelesini ya da bir türün diğeri için besin olduğu yırtıcı-av etkileşimini tanımlar. Diğer yazarların çalışmaları çeşitli modellerin inşasına odaklanmışsa, V. Volterra biyolojik toplulukların inşa edilmiş modelleri üzerinde derinlemesine bir çalışma yaptı. Birçok bilim adamına göre modern matematiksel ekoloji V. Volterra'nın kitabıyla başladı.2. TEMEL “YIRTICI-KURBAN” MODELİNİN NİTELİKSEL ARAŞTIRILMASI
2.1 “Yırtıcı-av” tipine göre trofik etkileşim modeli
V. Volterre tarafından oluşturulan "yırtıcı-av" tipi trofik etkileşim modelini ele alalım. Biri diğerini yiyen iki türden oluşan bir sistem olsun.
Türlerden birinin yırtıcı, diğerinin av olduğu durumu düşünün; yırtıcı hayvanın yalnızca avla beslendiğini varsayacağız. Aşağıdaki basit hipotezi kabul edelim:
- kurban büyüme hızı; - yırtıcı hayvan büyüme hızı; - avın popülasyon büyüklüğü; - yırtıcı hayvanın popülasyon büyüklüğü; - mağdurun doğal büyüme katsayısı; - yırtıcı hayvanın avını tüketme oranı; - avın yokluğunda yırtıcı hayvanın ölüm oranı; - avın biyokütlesinin avcı tarafından kendi biyokütlesine "işlenme" katsayısı.Daha sonra avcı-av sistemindeki popülasyon dinamikleri bir diferansiyel denklem sistemi (2.1) ile tanımlanacaktır:
(2.1)burada tüm katsayılar pozitif ve sabittir.
Modelin bir denge çözümü (2.2) vardır:
(2.2)Model (2.1)'e göre, yırtıcı hayvanların toplam hayvan kütlesindeki payı formül (2.3) ile ifade edilir:
(2.3)Küçük bozulmalara göre denge durumunun kararlılığının analizi, tekil noktanın (2.2) "nötr" kararlı ("merkez" tipinde) olduğunu, yani dengeden herhangi bir sapmanın ortadan kalkmadığını, ancak denge durumunu aktardığını göstermiştir. sistemi, bozukluğun büyüklüğüne bağlı olarak bir genliğe sahip bir salınım moduna geçirir. Faz düzlemindeki sistem yörüngeleri
denge noktasından çeşitli mesafelerde bulunan kapalı eğriler biçimindedir (Şekil 1).Pirinç. 1 – Klasik Volterra sistemi “yırtıcı-av”ın aşama “portresi”
Sistemin ilk denklemini (2.1) ikinciye bölerek faz düzlemindeki eğri için diferansiyel denklemi (2.4) elde ederiz.
. (2.4)Bu denklemi entegre edersek şunu elde ederiz:
(2.5) entegrasyon sabitidir, buradaBir noktanın faz düzlemi boyunca hareketinin yalnızca bir yönde gerçekleşeceğini göstermek kolaydır. Bunu yapmak için işlevleri değiştirmek uygundur
ve düzlemdeki koordinatların kökenini sabit bir noktaya (2.2) taşımak ve ardından kutupsal koordinatları eklemek: (2.6)Bu durumda sistem (2.6)'nın değerlerini sistem (2.1)'e koyarsak elimizde olur.
20'li yıllarda. A. Lotka ve bir süre sonra ondan bağımsız olarak V. Volter, yırtıcı ve av sayılarındaki eşlenik dalgalanmaları tanımlayan matematiksel modeller önerdi.
Model iki bileşenden oluşur:
C – yırtıcı hayvanların sayısı; N – mağdurların sayısı;
Yırtıcı hayvanların yokluğunda av popülasyonunun katlanarak büyüyeceğini varsayalım: dN/dt = rN. Ancak av, yırtıcı hayvan ile av arasındaki karşılaşma sıklığına göre belirlenen bir oranda yırtıcı hayvanlar tarafından yok edilir ve avcı (C) ve av (N) sayısı arttıkça buluşma sıklığı da artar. Karşılaşılan ve başarıyla yenen avın kesin sayısı, yırtıcı hayvanın avı bulma ve yakalama verimliliğine bağlı olacaktır; a' – “arama verimliliği” veya “saldırı sıklığı”. Böylece yırtıcı hayvan ile av arasındaki "başarılı" karşılaşmaların sıklığı ve dolayısıyla kurbanları yeme oranı a'СN'ye eşit olacaktır ve genel olarak: dN/dt = rN – a'CN (1*).
Yiyecek yokluğunda, bireysel yırtıcılar kilo verir, açlıktan ölür. Söz konusu modelde, açlık nedeniyle yiyecek yokluğunda yırtıcı hayvanın popülasyon büyüklüğünün katlanarak azalacağını varsayalım: dC/dt = - qC, burada q ölüm oranıdır. Ölüm, bu modelde iki duruma bağlı olduğuna inanılan bir oranda yeni bireylerin doğmasıyla telafi edilir:
1) gıda tüketim oranı, a’CN;
2) bu yiyeceğin yırtıcı hayvanın yavrularına geçme etkinliği (f).
Dolayısıyla avcının doğurganlığı fa'CN'ye eşittir ve genel olarak: dC/dt = fa'CN – qC (2*). Denklemler 1* ve 2* Lotka-Wolter modelini oluşturur. Bu modelin özellikleri incelenebilir, sabit bir popülasyon büyüklüğüne karşılık gelen çizgi izoklinleri oluşturulabilir ve bu tür izoklinlerin yardımıyla etkileşim halindeki yırtıcı-av popülasyonlarının davranışları belirlenebilir.
Av popülasyonu durumunda: dN/dt = 0, rN = a'CN veya C = r/a'. Çünkü r ve a’ = const, kurban için izoklin, C değerinin sabit olduğu doğru olacaktır:
Düşük avcı yoğunluğunda (C), av sayısı (N) artar, aksine azalır.
Benzer şekilde yırtıcı hayvanlar için (denklem 2*) dC/dt = 0, fa'CN = qC veya N = q/fa', yani. Yırtıcı hayvan için izoklin, N'nin sabit olduğu çizgi olacaktır: yüksek yoğunluk Av, avcının popülasyon büyüklüğü artar, azaldığında azalır.
Sayıları sınırsız eşlenik dalgalanmalara maruz kalır. Av sayısı fazla olduğunda avcıların sayısı artar, bu da avcıların av popülasyonu üzerindeki baskısının artmasına ve dolayısıyla sayısının azalmasına neden olur. Bu azalma, yırtıcı hayvanların yiyecek bakımından sınırlanmasına ve sayılarının azalmasına yol açar, bu da yırtıcı hayvanların baskısının zayıflamasına ve av sayısının artmasına neden olur, bu da yine yırtıcı hayvan popülasyonunun artmasına neden olur. , vesaire.
Popülasyonlar, bazı dış etkiler sayılarını değiştirene kadar uzun bir süre boyunca aynı salınım döngüsünü süresiz olarak gerçekleştirir, ardından popülasyonlar yeni sınırsız salınım döngüleri gerçekleştirir. Aslında çevre sürekli değişiyor ve nüfus büyüklükleri de sürekli değişiyor. yeni seviye. Bir popülasyonun yaptığı salınım döngülerinin düzenli olabilmesi için bunların istikrarlı olması gerekir: eğer bir dış etki popülasyon seviyesini değiştirirse, o zaman orijinal döngüye yönelmeleri gerekir. Bu tür çevrimlere kararlı, limit çevrimler denir.
Lotka-Wolter modeli, yırtıcı hayvan popülasyonundaki dalgalanmaların eşlik ettiği, av popülasyonundaki dalgalanmaların ortaya çıkmasıyla ifade edilen yırtıcı-av ilişkisindeki ana eğilimi göstermemize olanak tanır. Bu tür dalgalanmaların ana mekanizması, çok sayıda avdan çok sayıda yırtıcı hayvana, ardından az sayıda av ve az sayıda yırtıcı hayvana, ardından çok sayıda ava doğru giden durumlar dizisinin doğasında olan zaman gecikmesidir. vesaire.
5) YIRTICI VE PRİMATLARIN NÜFUS STRATEJİLERİ
“Yırtıcı-av” ilişkisi, madde ve enerjinin fitofajlardan zoofajlara veya alt düzey yırtıcılardan yırtıcılara aktarılması sürecindeki bağlantıları temsil eder. yüksek mertebeden. İle Bu ilişkilerin doğasına bağlı olarak üç tür yırtıcı ayırt edilir.:
A) toplayıcılar. Yırtıcı hayvan küçük, oldukça fazla sayıda hareketli kurban toplar. Bu tür bir yırtıcılık, enerjisini yalnızca kurban aramak için harcayan birçok kuş türü (cılıbıt, ispinoz, incir kuşu vb.) için tipiktir;
B) gerçek yırtıcılar. Yırtıcı hayvan avını takip edip öldürür;
V) pastoralistler. Bu avcılar, at sinekleri veya at sinekleri gibi avları tekrar tekrar kullanırlar.
Yırtıcı hayvanlardan yiyecek elde etme stratejisi, enerji verimliliği beslenme: Yiyecek elde etmek için harcanan enerji, asimilasyonu sırasında elde edilen enerjiden daha az olmalıdır.
Gerçek yırtıcılar bölünmüştür
Bol kaynaklarla (planktonik balıklar ve hatta balenli balinalar dahil) beslenen "orakçılar" ve daha az bol yiyecek arayan "avcılar". Sırasıyla
"Avcılar", avını bekleyen "pusucular" (örneğin turna, şahin, kedi, peygamber devesi böceği), "arayanlar" (böcek yiyen kuşlar) ve "takipçiler" olarak ayrılır. İkinci grup için, yiyecek aramak büyük miktarda enerji harcamayı gerektirmez, ancak avın (savanalardaki aslanlar) ele geçirilmesi için büyük miktarda enerjiye ihtiyaç vardır. Ancak bazı avcılar strateji unsurlarını birleştirebilir farklı seçenekler avcılık.
“Fitofaj-bitki” ilişkisinde olduğu gibi, tüm kurbanların avcılar tarafından yenildiği ve sonuçta ölüme yol açtığı bir durum doğada görülmez. Ekolojik denge Yırtıcı hayvanlar ve av arasındaki ilişki korunur özel mekanizmalar mağdurların tamamen yok olma riskini azaltır. Yani mağdurlar:
Bir yırtıcıdan kaçmak. Bu durumda, adaptasyonların bir sonucu olarak, hem kurbanların hem de yırtıcı hayvanların hareketliliği artar; bu, özellikle takipçilerinden saklanacak hiçbir yeri olmayan bozkır hayvanları için tipiktir;
Koruyucu renk elde edin (yaprak veya ince dal gibi davranın) veya tam tersi, parlak renk, N.: kırmızı, yırtıcıları acı tat konusunda uyarıyor. Bir tavşanın renginin değiştiği iyi bilinmektedir. farklı zamanlar yazın çimenlerde, kışın ise arka planda kendini kamufle etmesine olanak tanıyan yıl Beyaz kar. Uyarlanabilir renk değişiklikleri meydana gelebilir Farklı aşamalar birey oluşumu: fok yavruları beyazdır (kar rengi) ve yetişkinler siyahtır (kayalık sahil rengi);
Gruplar halinde dağıtın, bu da onları aramayı ve yakalamayı yırtıcı hayvan için daha enerji yoğun hale getirir;
Barınaklarda saklanın;
Aktif savunma önlemlerine geçin (boynuzlu otoburlar, dikenli balık), bazen eklem (misk öküzleri işgal edebilir " çok yönlü savunma"kurtlardan vb.).
Buna karşılık, yırtıcı hayvanlar yalnızca avını hızlı bir şekilde takip etme yeteneğini geliştirmekle kalmaz, aynı zamanda avın yerini kokuyla belirlemelerine olanak tanıyan bir koku alma duyusunu da geliştirir. Pek çok yırtıcı hayvan türü kurbanlarının (tilkiler, kurtlar) yuvalarını parçalar.
Aynı zamanda varlıklarının tespit edilmesini önlemek için kendileri de mümkün olan her şeyi yaparlar. Bu, kokuları gidermek için tuvalete çok fazla zaman harcayan ve dışkılarını gömen küçük kedilerin temizliğini açıklar. Yırtıcı hayvanlar “kamuflaj cüppeleri” giyerler (mızrak ve tüneklerden oluşan şeritler, onları makrofit çalılıkları, kaplan şeritleri vb. arasında daha az fark edilir hale getirir).
Yırtıcı hayvan popülasyonlarındaki tüm bireylerin yırtıcılardan tam olarak korunması da sağlanamaz, çünkü bu yalnızca açlıktan ölmek üzere olan yırtıcı hayvanların ölümüne yol açmakla kalmaz, aynı zamanda sonuçta av popülasyonlarının felaketine de yol açar. Aynı zamanda yırtıcı hayvanların popülasyon yoğunluğunun yokluğunda veya azalması durumunda, av popülasyonunun gen havuzu bozulur (hasta ve yaşlı hayvanlar alıkonulur) ve sayılarındaki keskin artış nedeniyle besin arzı zayıflar.
Bu nedenle, av ve avcıların popülasyon büyüklükleri arasındaki bağımlılığın etkisi - av popülasyonu büyüklüğünde bir dalgalanma, bunu takip eden yırtıcı popülasyon büyüklüğünde bir miktar gecikmeyle dalgalanma ("Lotka-Volterra etkisi") - nadiren gözlemlenir. .
Yırtıcı hayvanların ve avların biyokütlesi arasında oldukça istikrarlı bir oran kurulur. Böylece R. Ricklefs, yırtıcı ve av biyokütle oranının 1:150 ila 1:300 arasında değiştiğine dair veriler sağlar. Amerika Birleşik Devletleri'nin ılıman bölgesinin farklı ekosistemlerinde, bir kurt için 300 küçük beyaz kuyruklu geyik (ağırlık 60 kg), 100 büyük wapiti geyiği (ağırlık 300 kg) veya 30 geyik (ağırlık 350) vardır. Aynı model savanlarda da görüldü.
Bitkisel popülasyonların yoğun şekilde sömürülmesi nedeniyle insanlar genellikle avcıları ekosistemlerden dışlar (örneğin Büyük Britanya'da karaca ve geyik vardır, ancak kurt yoktur; sazan ve diğer havuz balıklarının yetiştirildiği yapay rezervuarlarda mızrak yoktur). Bu durumda, yırtıcı hayvanın rolü, fitofaj popülasyonunun bireylerinin bir kısmını ortadan kaldırarak kişinin kendisi tarafından gerçekleştirilir.
Bitkilerde ve mantarlarda özel bir yırtıcılık çeşidi gözlenir. Bitkiler aleminde böcekleri yakalayabilen ve onları proteolitik enzimlerin yardımıyla kısmen sindirebilen yaklaşık 500 tür vardır. Yırtıcı mantarlar miselyumun kısa dallarında bulunan küçük oval veya küresel başlıklar şeklinde yakalama cihazları oluşturur. Ancak en yaygın tuzak türü üç boyutlu yapışkan ağlardır. çok sayıda hiflerin dallanması sonucu oluşan halkalar. Yırtıcı mantarlar oldukça büyük hayvanları yakalayabilir; yuvarlak kurtlar. Solucan hiphalara dolandıktan sonra, hayvanın vücudunun içinde büyürler ve hızla onu doldururlar.
1. Sabit ve uygun sıcaklık ve nem seviyeleri.
2. Yiyecek bolluğu.
3.Olumsuz etkenlerden korunma.
4. Agresif kimyasal bileşim habitat (sindirim suları).
1. İki habitatın varlığı: Birinci derece ortam, konakçı organizmadır, ikinci derece ortam ise dış ortamdır.
PA88 sistemi, bir maddenin 100'den fazla farmakolojik etkisinin olasılığını ve etki mekanizmasını eş zamanlı olarak tahmin eder. yapısal formül. Bu yaklaşımın tarama planlamasındaki etkinliği yaklaşık %800'dür ve bilgisayar tahmininin doğruluğu uzmanların tahmininden %300 daha yüksektir.
Dolayısıyla tıpta yeni bilgi ve çözümler elde etmenin yapıcı araçlarından biri de matematiksel modelleme yöntemidir. Tıbbın matematikleştirilmesi süreci, iç içe geçmenin sık görülen bir tezahürüdür bilimsel bilgi, tedavi ve önleyici çalışmaların verimliliğini arttırmak.
4. “Yırtıcı-av” matematiksel modeli
Biyolojide ilk kez, İtalyan matematikçi V. Volterra ve meslektaşları tarafından, düşman hayvan türlerinin sayısındaki periyodik değişimlerin matematiksel modeli önerildi. Volterra'nın önerdiği model, 1924'te A. Lottka'nın “Fiziksel Biyolojinin Unsurları” kitabında ortaya attığı fikrin geliştirilmiş haliydi. Bu nedenle bu klasik matematiksel model “Lottky-Volterra” modeli olarak bilinmektedir.
Doğada antagonist türlerin ilişkileri bir modeldekinden daha karmaşık olmasına rağmen, bunlar yine de matematiksel modellemenin temel fikirlerinin çalışılacağı iyi bir öğretim modelidir.
Yani sorun şu: Ekolojik olarak kapalı bazı bölgelerde iki tür hayvan yaşıyor (örneğin vaşak ve tavşan). Yabani tavşan (av), her zaman yeterli miktarda mevcut olan bitki besinleriyle beslenir (bu model, sınırlı kaynakları hesaba katmaz) gübre). Vaşaklar (yırtıcı hayvanlar) yalnızca tavşan yiyebilir. Böyle bir durumda av ve yırtıcı hayvan sayısının zaman içinde nasıl değişeceğinin belirlenmesi gerekmektedir. ekolojik sistem. Av popülasyonu artarsa avcılarla avın karşılaşma olasılığı artar ve buna bağlı olarak belirli bir süre sonra avcı popülasyonu artar. Oldukça basit olan bu model, doğadaki gerçek yırtıcı hayvan popülasyonları ile av arasındaki etkileşimi oldukça yeterli bir şekilde açıklamaktadır.
Şimdi başlayalım diferansiyel denklemlerin hazırlanması. Hakkında
av sayısını N ile, yırtıcı hayvanların sayısını da M ile gösterelim. N ve M sayıları t süresinin fonksiyonlarıdır. Modelimizde aşağıdaki faktörleri dikkate alıyoruz:
a) mağdurların doğal üremesi; b) mağdurların doğal ölümü;
c) kurbanların yırtıcı hayvanlar tarafından yenilerek yok edilmesi; d) yırtıcı hayvanların doğal olarak yok olması;
e) Yiyecek varlığında üreme nedeniyle yırtıcı hayvanların sayısında artış.
Çünkü Hakkında konuşuyoruzÖ matematiksel model o zaman görev, amaçlanan tüm faktörleri içeren ve dinamikleri, yani yırtıcı hayvanların ve avların sayısındaki zaman içindeki değişimi açıklayan denklemler elde etmektir.
Av ve avcıların sayısının belirli bir t süresi boyunca ∆N ve ∆M kadar değişmesine izin verin. Kurban sayısındaki ∆N zaman içindeki ∆t değişimi, öncelikle doğal üremenin (mevcut kurban sayısıyla orantılı olan) bir sonucu olarak ortaya çıkan artışla belirlenir:
burada B, mağdurların doğal olarak yok olma oranını karakterize eden orantılılık katsayısıdır.
Yırtıcı hayvanlar tarafından tüketilmesi nedeniyle av sayısında azalmayı açıklayan denklemin türetilmesi, ne kadar sık karşılaşılırsa av sayısının o kadar hızlı azaldığı fikrine dayanmaktadır. Yırtıcı hayvanlarla av arasındaki karşılaşma sıklığının hem kurbanların hem de yırtıcıların sayısıyla orantılı olduğu da açıktır.
Denklemin (4) sol ve sağ taraflarını ∆t'ye bölerek ve ∆t→0'daki limite geçerek birinci dereceden bir diferansiyel denklem elde ederiz:
Bu denklemi çözebilmek için yırtıcı hayvan sayısının (M) zaman içinde nasıl değiştiğini bilmeniz gerekir. Yırtıcı hayvan sayısındaki değişiklik (∆M), yeterli gıdanın varlığında doğal üreme nedeniyle bir artış (M 1 = Q∙N∙M∙∆t) ve yırtıcı hayvanların doğal olarak yok olması nedeniyle bir azalma ile belirlenir ( M 2 = - P∙M∙∆ t):
M = Q∙N∙M∙∆t - P∙M∙∆t |
Denklem (6)'dan diferansiyel denklemi elde edebiliriz:
Diferansiyel denklemler (5) ve (7) matematiksel bir “yırtıcı-av” modelini temsil etmektedir. Katsayının değerlerini belirlemek yeterlidir

A, B, C, Q, P ve problemi çözmek için bir matematiksel model kullanılabilir.
Matematiksel modelin kontrol edilmesi ve ayarlanması. Bu laboratuvarda
En eksiksiz matematiksel modeli (denklem 5 ve 7) hesaplamanın yanı sıra, bir şeyin dikkate alınmadığı daha basit modellerin de incelenmesi önerilmektedir.
Matematiksel modelin beş karmaşıklık düzeyini göz önünde bulundurarak, modeli kontrol etme ve ayarlama aşamasını "hissedebilirsiniz".
1. seviye – modelde “avlar” için yalnızca doğal üremeleri dikkate alınır, “yırtıcı hayvanlar” yoktur;
Seviye 2 – model “avların” doğal yok oluşunu dikkate alır, “yırtıcı” yoktur;
Seviye 3 – model “kurbanların” doğal üremesini dikkate alır
Ve nesli tükeniyor, “yırtıcı hayvanlar” yok;
4. seviye – model “kurbanların” doğal üremesini hesaba katıyor
Ve neslinin tükenmesi ve "yırtıcı hayvanlar" tarafından yenilmesi, ancak "yırtıcı hayvanların" sayısı değişmeden kalır;
Seviye 5 – model tartışılan tüm faktörleri dikkate alır.
Böylece sahibiz aşağıdaki sistem diferansiyel denklemler:
M, “yırtıcıların” sayısıdır; N – “kurbanların” sayısı;
t – şimdiki zaman;
A – “kurbanların” üreme oranı; C – yırtıcı-av karşılaşmalarının sıklığı; B – “kurbanların” yok olma oranı;
Q – “yırtıcı hayvanların” üremesi;
P – “yırtıcı hayvanların” yok olması.
1. seviye: M = 0, B = 0; 2. seviye: M = 0, A = 0; 3. seviye: M = 0; 4. seviye: Q = 0, P = 0;
Seviye 5: tam denklem sistemi.
Katsayıların değerlerini her seviyeye değiştirerek farklı çözümler elde edeceğiz, örneğin:
3. seviye için M=0 katsayısının değeri, o zaman

elde ettiğimiz denklemi çözüyoruz
Aynı şekilde 1. ve 2. seviyeler için de. 4. ve 5. seviyelere gelince, burada denklem sistemini Runge-Kutta yöntemini kullanarak çözmek gerekiyor. Sonuç olarak bu seviyelerin matematiksel modellerine bir çözüm elde ediyoruz.
II. UYGULAMALI DERSLERDE ÖĞRENCİLERİN ÇALIŞMASI
1. Egzersiz . Sözlü konuşma kontrolü ve edinim düzeltmesi teorik materyal sınıflar. Derslere kabulü geçmek.
Görev 2. Laboratuvar çalışmasının yapılması, elde edilen sonuçların tartışılması, not yazılması.
İşin tamamlanması
1. Bilgisayar masaüstünden, farenin sol tuşuyla ilgili kısayola çift tıklayarak “Lab No. 6” programını çağırın.
2. "PREDATOR" kısayolunda farenin sol tuşuna çift tıklayın.
3. "PRED" kısayolunu seçin ve sol fare düğmesini kullanarak (çift tıklayarak) program çağrısını tekrarlayın.
4. Başlık ekranından sonra "ENTER" tuşuna basın.
5. Modelleme şununla başlar: 1. seviye.
6. Modelin analiz edileceği yılı girin: örneğin 2000
7. Zaman aralıklarını seçin; örneğin 40 yıl içinde, 1 yıl sonra (sonra 4 yıl sonra).
2. düzey: B = 0,05; N0 = 200;
3. seviye: A = 0,02; B = 0,05; N = 200;
4. düzey: A = 0,01; B = 0,002; C = 0,01; N0 = 200; M = 40; 5. seviye: A = 1; B = 0,5; C = 0,02; S = 0,002; P = 0,3; N0 = 200;
9. Denklemleri, grafikleri, modelin özelliklerinin hesaplanmasının sonuçlarını, yapılan işe ilişkin sonuçları içermesi gereken çalışma hakkında yazılı bir rapor hazırlayın.
Görev 3. Nihai bilgi seviyesinin izlenmesi:
a) Tamamlanan çalışmaya ilişkin sözlü rapor laboratuvar işi; b) durumsal sorunları çözmek; c) bilgisayar testi.
Görev 4. Bir sonraki ders için ödev: dersin bölümü ve konusu, özet raporlar için konuların koordinasyonu (raporun uzunluğu 2-3 sayfa, süre sınırı 5-7 dakika).
Yırtıcılar otçulları ve ayrıca zayıf yırtıcıları yiyebilirler. Yırtıcı hayvanların geniş bir yiyecek yelpazesi vardır ve bir avdan diğerine, daha erişilebilir olana kolayca geçiş yaparlar. Yırtıcı hayvanlar genellikle zayıf avlara saldırır. Av-yırtıcı popülasyonları arasında ekolojik denge korunur.[...]
Denge kararsızsa (limit döngüler yoksa) veya dış döngü kararsızsa, o zaman güçlü dalgalanmalar yaşayan her iki türün sayısı dengenin yakınını terk eder. Ayrıca, yırtıcı hayvanın adaptasyonunun düşük olması durumunda hızlı dejenerasyon (ilk durumda) meydana gelir, yani. yüksek ölüm oranıyla (kurbanın üreme hızıyla karşılaştırıldığında). Bu, her bakımdan zayıf olan bir yırtıcının, sistemin istikrarına katkıda bulunmadığı ve kendisinin yok olduğu anlamına gelir.[...]
Avcı-av birlikte evriminde denge avcıya doğru kaydığında ve avın menzili daraldığında yırtıcı hayvanların baskısı özellikle güçlü olur. Rekabetçi mücadele, gıda kaynaklarının eksikliğiyle yakından ilişkilidir; örneğin, yırtıcıların kaynak olarak uzaya yönelik doğrudan bir mücadelesi de olabilir, ancak çoğu zaman bu, yeterli yiyeceğe sahip olmayan bir türün yer değiştirmesidir. Aynı miktarda yiyeceğe sahip bir türe verilen bölge. Çoktan türler arası rekabet.[ ...]
 |
Ve son olarak, model (2.7) tarafından açıklanan "yırtıcı-av" sisteminde, yayılma istikrarsızlığının ortaya çıkması (yerel denge kararlılığı ile), ancak yırtıcı hayvanın doğal ölüm oranının, popülasyonunun normalden daha hızlı artmasıyla birlikte artması durumunda mümkündür. doğrusal fonksiyon ve trofik fonksiyon Volterra'nınkinden farklıdır veya av popülasyonu Ollie tipi bir popülasyondur [...]
Teorik olarak, "bir yırtıcı - iki av" modelinde eşdeğer otlatma (bir veya başka bir av türü için tercih eksikliği), yalnızca potansiyel olarak istikrarlı bir dengenin zaten mevcut olduğu yerlerde av türlerinin rekabetçi bir arada varoluşunu etkileyebilir. Çeşitlilik ancak rekabet kabiliyeti daha az olan türlerin, baskın türlere göre daha yüksek nüfus artış hızına sahip olduğu koşullar altında artabilir. Bu, tekdüze otlatmanın bitki türü çeşitliliğinde artışa yol açtığı durumu anlamamızı sağlar. daha büyük sayı Hızlı üreme için seçilen türler, evrimi rekabet gücünü artırmayı amaçlayan türlerle bir arada bulunur.[...]
Benzer şekilde, yoğunluğa bağlı av seçimi, daha önce hiçbir dengenin bulunmadığı iki rakip av türünün teorik modellerinde istikrarlı bir dengeye yol açabilir. Bunu yapabilmek için yırtıcı hayvanın, av yoğunluğundaki değişikliklere fonksiyonel ve sayısal tepkiler verme yeteneğine sahip olması gerekir; Ancak değişimin (en bol ava orantısız sıklıkta yapılan saldırılar) daha fazla etki yaratması mümkündür. önemli. Aslında, geçişin "bir yırtıcı - n av" sistemlerinde dengeleyici bir etkiye sahip olduğu ve av nişlerinin tamamen örtüştüğü durumlarda etkileşimleri dengeleyebilen tek mekanizmayı temsil ettiği tespit edilmiştir. Uzmanlaşmamış avcılar böyle bir rol oynayabilir. Daha uzmanlaşmış avcıların baskın bir rakip için tercihi, avcının değişmesiyle aynı şekilde hareket eder ve nişlerinin bir dereceye kadar ayrılmış olması koşuluyla, av türleri arasında daha önce hiçbir dengenin bulunmadığı modellerde teorik etkileşimleri dengeleyebilir.[...]
Ayrıca, "her bakımdan güçlü" bir yırtıcı, topluluğu istikrara kavuşturmaz; belirli bir ava iyi adapte olmuş ve göreceli ölüm oranı düşük. Bu durumda sistem kararsız bir sınır döngüsüne sahiptir ve denge pozisyonunun stabilitesine rağmen rastgele bir ortamda dejenere olur (yırtıcı hayvan avını yer ve sonuç olarak ölür). Bu durum yavaş dejenerasyona karşılık gelir.[...]
Bu nedenle, yırtıcı hayvanın istikrarlı bir dengeye iyi bir şekilde uyum sağlamasıyla, istikrarsız ve istikrarlı döngüler ortaya çıkabilir; bağlı olarak başlangıç koşulları“yırtıcı-av” sistemi ya dengeye yönelir ya da salınarak ondan uzaklaşır ya da denge yakınında her iki türün sayısında istikrarlı dalgalanmalar oluşur [...]
Yırtıcı olarak sınıflandırılan organizmalar diğer organizmalarla beslenerek avlarını yok ederler. Bu nedenle, canlı organizmalar arasında "yırtıcı hayvanlar" ve "av" olmak üzere bir sınıflandırma sistemi daha ayırt edilmelidir. Bu tür organizmalar arasındaki ilişkiler, gezegenimizdeki yaşamın evrimi boyunca gelişmiştir. Yırtıcı organizmalar, av organizmalarının sayısının doğal düzenleyicileri olarak görev yapar. "Yırtıcı hayvanların" sayısındaki bir artış, "avların" sayısında bir azalmaya yol açar, bu da "yırtıcı hayvanlar" için yiyecek ("av") tedarikini azaltır, bu da genellikle sayının azalmasını gerektirir. Böylece biyosinozda yırtıcı ve av sayısında sürekli dalgalanmalar meydana gelir, ancak genel olarak belirli bir süre boyunca adil bir denge içinde belirli bir denge kurulur. istikrarlı koşullarçevre.[...]
Bu sonuçta yırtıcı ve av popülasyonları arasında ekolojik bir dengeye ulaşıyor.[...]
Üçüncü tipteki bir trofik fonksiyon için, N fonksiyonun bükülme noktası olduğunda denge durumu stabil olacaktır (bkz. Şekil 2, c). Bu, aralık boyunca trofik fonksiyonun içbükey olması ve dolayısıyla avcının av tüketiminin göreceli payının artması gerçeğinden kaynaklanmaktadır.[...]
Гг = -Г olsun, yani. “yırtıcı-avcı” tipi bir topluluk var. Bu durumda (7.4) numaralı ifadedeki ilk terim sıfıra eşit olup, N denge durumunun olasılığı açısından kararlılık koşulunu sağlamak için ikinci terimin de pozitif olmaması gerekir.[.. .]
Dolayısıyla, ele alınan yırtıcı-av topluluğu için, genel pozitif denge konumunun asimptotik olarak kararlı olduğu sonucuna varabiliriz, yani herhangi bir 1H(0)>0 başlangıç verisi için evrim, N(7) - ■ K olacak şekilde gerçekleşir. N >0 olması koşuluyla.[...]
Böylece üreme için barınağı olmayan homojen bir ortamda, yırtıcı hayvan er ya da geç avın popülasyonunu yok eder ve sonra kendi kendine yok olur. Yaşam dalgaları” (yırtıcı hayvanın ve avın bolluğundaki değişiklikler) sürekli bir faz değişimiyle birbirini takip eder ve ortalama olarak hem avcının hem de avın bolluğu yaklaşık olarak aynı seviyede kalır. Dönemin süresi her iki türün büyüme oranlarına ve başlangıç parametrelerine bağlıdır. Av popülasyonu için yırtıcı hayvanın etkisi olumludur çünkü aşırı üremesi popülasyonunda bir çöküşe yol açacaktır. Buna karşılık, avın tamamen yok edilmesini engelleyen tüm mekanizmalar, yırtıcı hayvanın yiyecek tedarikinin korunmasına katkıda bulunur.[...]
Diğer değişiklikler yırtıcı hayvanın davranışının bir sonucu olabilir. Bir yırtıcı hayvanın tüketebileceği av birey sayısı verilen zaman, bir sınırı var. Bu eşiğe yaklaşıldığında yırtıcı hayvan doygunluğunun etkisi Tablo'da gösterilmektedir. 2-4, B. Denklem 5 ve 6 ile açıklanan etkileşimler kararlı denge noktalarına sahip olabilir veya döngüsel dalgalanmalar sergileyebilir. Ancak bu tür döngüler, Lotka-Volterra denklemleri 1 ve 2'de yansıtılanlardan farklıdır. Denklemler 5 ve 6 tarafından aktarılan döngüler, ortam sabit olduğu sürece sabit genliğe ve ortalama yoğunluğa sahip olabilir; bir bozulma meydana geldikten sonra önceki genliklerine ve ortalama yoğunluklarına dönebilirler. Bozulmalardan kurtulan bu tür çevrimlere kararlı limit çevrimleri denir. Bir tavşan ile vaşak arasındaki etkileşim, kararlı bir sınır döngüsü olarak düşünülebilir, ancak bu bir Lotka-Volterra döngüsü değildir.[...]
Yırtıcı-av sisteminde difüzyon kararsızlığının oluşmasını ele alalım, ancak önce n = 2 olan (1.1) sisteminde difüzyon kararsızlığının oluşmasını sağlayan koşulları yazacağız. (N, N) dengesinin olduğu açıktır. ) yereldir (yani[ .. .]
Yırtıcı ve avın uzun süreli bir arada yaşamasıyla ilgili vakaların yorumlanmasına geçelim. Limit döngülerin yokluğunda, rastgele bir ortamda istikrarlı bir dengenin popülasyon dalgalanmalarına karşılık geleceği ve bunların genliğinin, bozuklukların dağılımıyla orantılı olacağı açıktır. Bu fenomen, yırtıcı hayvanın göreceli ölüm oranı yüksekse ve aynı zamanda belirli bir ava yüksek derecede uyum sağlama yeteneği varsa ortaya çıkacaktır.[...]
Şimdi avcının uygunluğunun artmasıyla sistem dinamiğinin nasıl değiştiğini ele alalım. b 1'den 0'a azalıyor. Eğer uygunluk yeterince düşükse, o zaman limit çevrimleri yoktur ve denge kararsızdır. Bu denge civarında uygunluktaki bir artışla birlikte, önce istikrarlı bir döngü, sonra da dışsal kararsız bir döngü ortaya çıkabilir. Başlangıç koşullarına bağlı olarak (avcı ve av biyokütlesi oranı), sistem ya stabilitesini kaybedebilir; dengenin yakınını terk edin, aksi takdirde zamanla içinde istikrarlı salınımlar oluşacaktır. Uygunluktaki daha fazla artış, sistem davranışının salınımlı doğasını imkansız hale getirir. Ancak ne zaman b [...]
Negatif (stabilize edici) geri bildirime bir örnek, avcı ile av arasındaki ilişki veya okyanus karbonat sisteminin işleyişidir (sudaki CO2 çözeltisi: CO2 + H2O -> H2CO3). Tipik olarak okyanus suyunda çözünmüş karbondioksit miktarı, atmosferdeki karbondioksit konsantrasyonuyla kısmi dengededir. Volkanik patlamalardan sonra atmosferdeki karbondioksitteki yerel artışlar, fotosentezin yoğunlaşmasına ve bunun okyanus karbonat sistemi tarafından emilmesine yol açar. Atmosferdeki karbondioksit seviyeleri azaldıkça okyanus karbonat sistemi atmosfere CO2 salıyor. Bu nedenle atmosferdeki karbondioksit konsantrasyonu oldukça stabildir.[...]
[ ...]
R. Ricklefs'in (1979) belirttiği gibi, “avcı-avcı” sisteminde ilişkilerin istikrara kavuşturulmasına katkıda bulunan faktörler vardır: Yırtıcının verimsizliği, yırtıcı için alternatif yiyecek kaynaklarının bulunması, avlanmadaki gecikmenin azaltılması. yırtıcı hayvanın tepkisi ve uygulanan çevresel kısıtlamalar dış ortam bir nüfus veya diğeri için. Yırtıcı ve av popülasyonları arasındaki etkileşimler oldukça çeşitli ve karmaşıktır. Böylece avcılar yeterince verimliyse avın nüfus yoğunluğunu düzenleyerek ortamın taşıma kapasitesinin altında tutabilirler. Yırtıcı hayvanlar, av popülasyonları üzerindeki etkileri aracılığıyla avın çeşitli özelliklerinin evrimini etkiler ve bu da sonuçta yırtıcı ve av popülasyonları arasında ekolojik dengenin kurulmasına yol açar.[...]
Koşullardan biri karşılanırsa: 0 1/2. Eğer 6 > 1 (kA [...]
Biyotanın ve çevrenin stabilitesi yalnızca bitkilerin (ototroflar ve otçul heterotrofik organizmalar) etkileşimine bağlıdır. Büyüklüğü ne olursa olsun yırtıcılar topluluğun ekolojik dengesini bozamazlar. doğal şartlar Kurban sayısı sabit kalırsa sayılarını artıramazlar. Yırtıcı hayvanların yalnızca hareketli olmaları gerekmez, aynı zamanda yalnızca hareketli hayvanlarla da beslenebilirler.[...]
Turna balığı kadar yaygın olan başka bir balık yoktur. Duran veya akan rezervuarlardaki birkaç balıkçılık alanında, av ve yırtıcı hayvan arasındaki dengeyi korumak için mızraklardan kaynaklanan bir baskı yoktur. Yalnızca, diğer balıkların üremesi nedeniyle turna balıklarının istenmeyen balıklar olduğu modern yapay rezervuarlar, bilinçli olarak iskan edilmemektedir. onlara. Pike dünyada son derece iyi temsil edilmektedir. Amerika Birleşik Devletleri ve Kanada'dan kuzey yarımkürede yakalanırlar. Kuzey Amerika, Avrupa üzerinden Kuzey Asya'ya.[...]
Sürdürülebilir bir arada yaşamanın başka bir olasılığı burada, nispeten yüksek uyumun dar bir aralığında ortaya çıkıyor. Çok "iyi" bir yırtıcının olduğu istikrarsız bir rejime geçiş sırasında, biyokütle dağılımının sisteme girişiyle (avın yüksek üretkenliği) dengelendiği istikrarlı bir dış sınır döngüsü ortaya çıkabilir. Daha sonra en muhtemel ikisinin olduğu ilginç bir durum ortaya çıkıyor karakteristik değerler rastgele salınımların genlikleri. Bazıları dengeye yakın, diğerleri ise sınır döngüsüne yakın meydana gelir ve bu rejimler arasında daha fazla veya daha az sıklıkta geçişler mümkündür [...]
Şekil 2'deki vektörlere göre davranan varsayımsal popülasyonlar. 10.11 A, Şekil 1'de gösterilmektedir. Şekil 10.11,-B'de yırtıcı ve av popülasyonlarının oranının dinamiklerini gösteren bir grafik kullanılmıştır. 10.11.5 yırtıcı hayvan ve av bolluğunun zaman içindeki dinamiklerini gösteren bir grafik şeklinde. Av popülasyonunda, düşük yoğunluklu bir dengeden yüksek yoğunluklu bir dengeye geçip geri döndüğünde, bir sayı "patlaması" meydana gelir. Ve sayılardaki bu artış, aynı derecede belirgin bir değişimin sonucu değil. çevre. Tam tersine, sayılardaki bu değişiklik etkinin kendisinden kaynaklanmaktadır (ortamdaki küçük bir “gürültü” seviyesi ile) ve özellikle çeşitli denge durumlarının varlığını yansıtmaktadır. Daha fazlasını açıklamak için benzer akıl yürütme kullanılabilir karmaşık vakalar Doğal popülasyonlarda popülasyon dinamikleri.[...]
Bir ekosistemin en önemli özelliği istikrarı, değişim dengesi ve içinde meydana gelen süreçlerdir. Popülasyonların veya ekosistemlerin değişen çevre koşullarında istikrarlı dinamik dengeyi sürdürme yeteneğine homeostazis (homoios - aynı, benzer; durağanlık - durum) denir. Homeostazis geri bildirim ilkesine dayanır. Doğada dengeyi korumak gerektirmez Harici kontrol. Homeostazisin bir örneği, yırtıcı ve avın popülasyon yoğunluğunun düzenlendiği "yırtıcı-av" alt sistemidir.[...]
Doğal ekosistem(biyojeosinoz), elementlerinin sürekli etkileşimi, maddelerin dolaşımı, kimyasal, enerjik, genetik ve diğer enerji ve bilgilerin zincirler-kanallar yoluyla aktarımı ile istikrarlı bir şekilde çalışır. Denge ilkesine göre, içinden enerji ve bilgi akışı geçen herhangi bir doğal sistem, kararlı bir durum geliştirme eğilimindedir. Aynı zamanda ekosistemlerin istikrarı bir geri bildirim mekanizması aracılığıyla otomatik olarak sağlanır. Geri bildirim, ekosistemlerin yönetilen bileşenlerinden alınan verilerin, yönetim bileşenleri tarafından süreçte ayarlamalar yapmak için kullanılmasından oluşur. Yukarıda bu bağlamda ele alınan “yırtıcı”-”av” ilişkisini biraz daha detaylı anlatmak gerekirse; evet, su ekosisteminde yırtıcı balık(havuzda turna balığı) diğer av balık türlerini (havuz sazanı) yiyin; havuz sazanlarının sayısı artarsa bu olumlu bir geri bildirim örneğidir; Havuz sazanı ile beslenen turna balığı sayılarını azaltır - bu olumsuz geri bildirimin bir örneğidir; Yırtıcı hayvanların sayısı arttıkça kurbanların sayısı azalır ve yiyecek kıtlığı yaşayan yırtıcı hayvan da popülasyonunun artışını azaltır; Sonuçta söz konusu gölette hem turna balığı hem de havuz sazanı sayısında dinamik bir denge kurulur. Denge sürekli olarak korunur, bu da herhangi bir bağlantının kaybolmasını önleyecektir trofik zincir(Şekil 64).[...]
Şimdi en önemli genellemeye geçelim: Ekosistem yeterince istikrarlı ve istikrarlıysa olumsuz etkileşimler zamanla daha az fark edilir hale gelir. mekânsal yapı Popülasyonların karşılıklı adaptasyonuna olanak sağlar. Gibi model sistemlerde yırtıcı-av Lotka-Volterra denklemiyle açıklanan, eğer denkleme sayıların kendi kendini sınırlama faktörlerinin eylemini karakterize eden ek terimler eklenmezse, o zaman salınımlar sürekli olarak meydana gelir ve kaybolmaz (bkz. Lewontin, 1969). Pimentel (1968; ayrıca bkz. Pimentel ve Stone, 1968) deneysel olarak bu tür ek terimlerin karşılıklı adaptasyonları veya genetik geri bildirimi yansıtabileceğini gösterdi. Sayılarının önemli dalgalanmalara maruz kaldığı bir kültürde daha önce iki yıl boyunca bir arada yaşayan bireylerden yeni kültürler yaratıldığında, popülasyonların her birinin diğeri tarafından "bastırıldığı" ekolojik homeostazı geliştirdikleri ortaya çıktı. öyle ki, daha istikrarlı bir dengede bir arada yaşamaları mümkün hale geldi.
İki türün etkileşim modelleri
Volterra'nın hipotezleri. Kimyasal kinetik ile analojiler. Volterra etkileşim modelleri. Etkileşim türlerinin sınıflandırılması Rekabet. Yırtıcı-av. Tür etkileşimlerinin genelleştirilmiş modelleri . Kolmogorov'un modeli. MacArthur'un iki böcek türü arasındaki etkileşim modeli. Parametrik ve Bazykin sisteminin faz portreleri.
Modern matematiksel popülasyon teorisinin kurucusu, aparatı diferansiyel ve integro-diferansiyel denklemler olan biyolojik toplulukların matematiksel teorisini geliştiren İtalyan matematikçi Vito Volterra olarak kabul edilir.(Vito Volterra. Lecons sur la Theorie Mathematique de la Lutte pour la Vie. Paris, 1931). Sonraki yıllarda nüfus dinamikleri esas olarak bu kitapta ifade edilen fikirler doğrultusunda gelişti. Volterra’nın kitabının Rusça çevirisi 1976 yılında “Varoluş Mücadelesinin Matematiksel Teorisi” başlığıyla Yu.M. Svirezhev, 1931–1976 döneminde matematiksel ekolojinin gelişim tarihini inceliyor.
Volterra'nın kitabı matematik kitaplarının yazıldığı şekilde yazılmıştır. Öncelikle üzerinde çalışılması gereken matematiksel nesneler hakkında bazı varsayımlar formüle eder ve daha sonra bu nesnelerin özelliklerine ilişkin matematiksel bir çalışma yürütür.
Volterra'nın incelediği sistemler iki veya daha fazla türden oluşur. Bazı durumlarda kullanılan gıdanın tedariki dikkate alınır. Bu türlerin etkileşimini açıklayan denklemler aşağıdaki kavramlara dayanmaktadır.
Volterra'nın hipotezleri
1. Yiyecek ya sınırsız miktarda bulunur ya da tedariki zaman içinde sıkı bir şekilde düzenlenir.
2. Her türün bireyleri öyle bir şekilde ölür ki, birim zamanda mevcut bireylerin sabit bir oranı ölür.
3. Yırtıcı türler kurbanları yerler ve birim zaman başına yenen kurbanların sayısı her zaman bu iki türün bireyleriyle karşılaşma olasılığıyla orantılıdır, yani. Yırtıcı hayvanların sayısı ile av sayısının çarpımı.
4. Sınırlı miktarda yiyecek varsa ve onu tüketebilecek birden fazla tür varsa, o zaman bir türün birim zamanda tükettiği yiyeceğin payı, bu türün birey sayısıyla orantılı olup, belirli bir katsayı ile alınır. türler (spesifik rekabet modelleri).
5. Bir tür sınırsız miktarda bulunan besinlerle besleniyorsa birim zamandaki tür sayısındaki artış, tür sayısıyla orantılıdır.
6. Bir türün sınırlı miktarlarda bulunan yiyeceklerle beslenmesi durumunda, bu türlerin üremesi, besin tüketim hızına göre düzenlenir; birim zaman başına artış, yenen yiyecek miktarıyla orantılıdır.
Kimyasal kinetik ile analojiler
Bu hipotezlerin kimyasal kinetikle yakın paralellikleri vardır. Popülasyon dinamiği denklemlerinde, kimyasal kinetik denklemlerinde olduğu gibi, reaksiyon hızı, reaksiyona giren bileşenlerin konsantrasyonlarının çarpımı ile orantılı olduğunda "çarpışma ilkesi" kullanılır.
Aslında Volterra'nın hipotezlerine göre hız işlem Her türün neslinin tükenmesi türün sayısıyla orantılıdır. Kimyasal kinetikte bu, belirli bir maddenin ayrışmasının monomoleküler reaksiyonuna karşılık gelir ve matematiksel modelde denklemlerin sağ taraflarındaki negatif doğrusal terimlere karşılık gelir.
Kimyasal kinetik kavramlarına göre, iki madde arasındaki etkileşimin bimoleküler reaksiyonunun hızı, bu maddelerin çarpışma olasılığı ile orantılıdır; konsantrasyonlarının ürünüdür. Aynı şekilde Volterra'nın hipotezlerine göre yırtıcı hayvanların üreme hızı (avın ölümü), yırtıcı ve av bireyleri arasındaki karşılaşma olasılığıyla orantılıdır; sayıların çarpımı. Her iki durumda da, model sisteminde karşılık gelen denklemlerin sağ tarafında çift doğrusal terimler görünür.
Son olarak, Volterra denklemlerinin sağ tarafındaki sınırsız koşullar altında popülasyonların büyümesine karşılık gelen doğrusal pozitif terimler, otokatalitik terimlere karşılık gelir. kimyasal reaksiyonlar. Kimyasal ve çevresel modellerdeki denklemlerin bu benzerliği, kimyasal reaksiyon sistemleri için olduğu gibi popülasyon kinetiğinin matematiksel modellemesi için aynı araştırma yöntemlerini uygulamamıza olanak tanır.
Etkileşim türlerinin sınıflandırılması
Volterra'nın hipotezlerine göre sayıları iki türün etkileşimi X 1 ve X 2 denklemlerle açıklanabilir:
(9.1)
İşte parametreler A Ben - türün kendi büyüme hızının sabitleri, C Ben- sayıların kendi kendini sınırlama sabitleri (spesifik rekabet), b ij- tür etkileşimi sabitleri, (Ben, j= 1,2). Bu katsayıların işaretleri etkileşimin türünü belirler.
Biyoloji literatüründe etkileşimler genellikle ilgili mekanizmalara göre sınıflandırılır. Buradaki çeşitlilik çok büyüktür: çeşitli trofik etkileşimler, bakteriler ve bakteriler arasında var olan kimyasal etkileşimler. planktonik algler mantarların diğer organizmalarla etkileşimi, bitki organizmalarının dizilişi, özellikle güneş ışığı rekabeti ve toprağın gelişimi vb. Bu sınıflandırma çok geniş görünüyor.
e . Odum, V. Volterra'nın önerdiği modelleri dikkate alarak mekanizmalara göre değil sonuçlara göre bir sınıflandırma önerdi. Bu sınıflandırmaya göre, bir türün bolluğunun başka bir türün varlığında artması, azalması veya değişmeden kalmasına bağlı olarak ilişkiler olumlu, olumsuz veya nötr olarak değerlendirilmelidir. Daha sonra ana etkileşim türleri tablo biçiminde sunulabilir.
TÜRLERİN ETKİLEŞİM TÜRLERİ
|
simbiyoz |
B 12 ,B 21 >0 |
||
|
KOMENSALİZM |
B 12 ,>0, B 21 =0 |
||
|
YIRTICI-KURBAN |
B 12 ,>0, B 21 <0 |
||
|
AMENSALİZM |
B 12 ,=0, B 21 <0 |
||
|
YARIŞMA |
B 12 , B 21 <0 |
||
|
TARAFSIZLIK |
B 12 , B 21 =0 |
Son sütun sistem (9.1)'den gelen etkileşim katsayılarının işaretlerini gösterir.
Ana etkileşim türlerine bakalım
REKABET DENKLEMLERİ:
Ders 6'da gördüğümüz gibi rekabet denklemleri:
 (9.2)
(9.2)
Sabit sistem çözümleri:
(1).
![]()
Herhangi bir sistem parametresi için koordinatların kökeni kararsız bir düğümdür.
(2).
![]() (9.3)
(9.3)
C durağan durum (9.3) bir eyerdir A 1 >b 12 /İle 2 ve
kararlı düğüm A 1 12 /s 2 . Bu durum, bir türün kendi büyüme hızının belirli bir kritik değerin altında olması durumunda neslinin tükenmesi anlamına gelir.
(3).
![]() (9.4)
(9.4)
C sabit çözüm (9.4)¾ eyerlenmek A 2 >b 21 /C 1 ve kararlı bir düğüm A 2< B 21 /C 1
(4).
![]() (9.5)
(9.5)
Durağan durum (9.5), iki rakip türün bir arada varlığını karakterize eder ve eğer ilişki sağlanırsa istikrarlı bir düğümü temsil eder:
![]()
Bu eşitsizliği ima eder:
B 12
B 21
türlerin bir arada yaşamasının koşulunu formüle etmemizi sağlar:
Popülasyonlar arası etkileşim katsayılarının çarpımı, popülasyon etkileşimi içindeki katsayıların çarpımından daha azdır.
Aslında, ele alınan iki türün doğal büyüme oranlarına izin verin.A 1 ,A 2 aynıdır. O zaman istikrar için gerekli koşul şu olacaktır:
C 2 > B 12 ,C 1 > b 21 .
Bu eşitsizlikler, bir rakibin büyüklüğündeki artışın, diğer bir rakibin büyümesine göre kendi büyümesini daha fazla bastırdığını göstermektedir. Her iki türün sayısının farklı kaynaklarla kısmen veya tamamen sınırlandırılması durumunda yukarıdaki eşitsizlikler geçerlidir. Her iki türün de tamamen aynı ihtiyaçları varsa, o zaman biri daha hayatta kalabilir ve rakibinin yerini alabilir.
Sistemin aşama yörüngelerinin davranışı, rekabetin olası sonuçları hakkında net bir fikir verir. Sistem (9.2) denklemlerinin sağ taraflarını sıfıra eşitleyelim:
X 1 (A 1 –c 1 X 1 – B 12 X 2) = 0 (dx 1 /dt = 0),
X 2 (A 2 –B 21 X 1 – C 2 X 2) = 0 (dx 2 /dt = 0),
Bu durumda sistemin ana izoklinleri için denklemler elde ederiz.
X 2 = – b 21 X 1 / C 2 +A 2/c2, X 2 = 0
– düşey teğetlerin izoklin denklemleri.
X 2 = –c1 X 1 / b 12 + A 1 /B 12 , X 1 = 0
– düşey teğetlerin izoklin denklemleri. Dikey ve yatay teğet sistemlerin izoklinlerinin ikili kesişme noktaları, denklem sisteminin (9.2.) durağan çözümlerini ve bunların koordinatlarını temsil eder. ![]() rekabet eden türlerin sabit sayılarıdır.
rekabet eden türlerin sabit sayılarıdır.
Sistemdeki (9.2) ana izoklinlerin olası konumu Şekil 9.1'de gösterilmektedir. Pirinç. 9.1Atürün hayatta kalmasına karşılık gelirX 1, şek. 9.1 B– türün hayatta kalmasıX 2, şek. 9.1 V– Koşul (9.6) karşılandığında türlerin bir arada bulunması. Şekil 9.1Gtetikleme sistemini gösterir. Burada rekabetin sonucu başlangıç koşullarına bağlıdır. Her iki tip için de sıfırdan farklı durağan durum (9.5) kararsızdır. Bu, her türün hayatta kalma alanlarını ayıran ayırıcının içinden geçtiği eyerdir.

Pirinç. 9.1.Farklı parametre oranlarına sahip iki tip (9.2) rekabetli Volterra sisteminin faz portresinde ana izoklinlerin konumu. Metindeki açıklamalar.
Tür rekabetini incelemek için çok çeşitli organizmalar üzerinde deneyler yapıldı. Tipik olarak birbirine yakın iki tür seçilir ve sıkı kontrol edilen koşullar altında birlikte ve ayrı ayrı büyütülür. Belirli aralıklarla nüfusun tam veya seçici bir sayımı yapılır. Birkaç tekrarlı deneyden elde edilen veriler kaydedilir ve analiz edilir. Protozoa (özellikle siliatlar), Tribolium cinsinin birçok böcek türü, drosophila ve tatlı su kabukluları (daphnia) üzerinde çalışmalar yapılmıştır. Mikrobiyal popülasyonlar üzerinde birçok deney yapılmıştır (bkz. ders 11). Planaryanlar (Reynolds), iki karınca türü (Pontin) vb. dahil olmak üzere doğada da deneyler yapıldı. 9.2. aynı kaynağı kullanan (aynı ekolojik nişi işgal eden) diatomların büyüme eğrilerini gösterir. Monokültürde yetiştirildiğinde Asterionella Formosa sabit bir yoğunluk seviyesine ulaşır ve kaynağın (silikat) konsantrasyonunu sürekli düşük bir seviyede tutar. B. Monokültürde yetiştirildiğinde Synedrauina benzer şekilde davranır ve silikat konsantrasyonunu daha da düşük bir seviyede tutar. B. Birlikte yetiştirme sırasında (iki kopya halinde) Synedrauina, Asterionella formosa'nın yerini alıyor. Görünüşe göre Synedra

Pirinç. 9.2.Diatomlarda rekabet. A - monokültürde yetiştirildiğinde Asterionella Formosa sabit bir yoğunluk seviyesine ulaşır ve kaynağın (silikat) konsantrasyonunu sürekli düşük bir seviyede tutar. B - monokültürde yetiştirildiğinde Synedrauina benzer şekilde davranır ve silikat konsantrasyonunu daha da düşük bir seviyede tutar. V- ortak yetiştirme ile (iki kopya halinde) Synedruina, Asterionella formosa'nın yerini alıyor. Görünüşe göre Synedra alt tabakayı daha iyi kullanma yeteneği nedeniyle rekabeti kazanır (ayrıca bkz. Ders 11).
G. Gause'un rekabeti incelemeye yönelik deneyleri yaygın olarak biliniyor; bu deneyler, rakip türlerden birinin hayatta kaldığını gösteriyor ve onun "rekabetçi dışlanma yasasını" formüle etmesine olanak tanıyor. Yasa, bir ekolojik nişte yalnızca bir türün var olabileceğini belirtiyor. İncirde. 9.3. Gause'un deneylerinin sonuçları, aynı ekolojik nişi işgal eden iki Parametium türü (Şekil 9.3 a, b) ve farklı ekolojik nişleri işgal eden türler (Şekil 9.3 c) için sunulmaktadır.

Pirinç. 9.3. A- İki türün nüfus artış eğrileri Parametyum tek tür bitkilerde. Siyah daireler – P Aurelia, beyaz daireler – P. kaudatum
B- P Aurelia ve P'nin büyüme eğrileri. kaudatum karma bir kültürde.
Gause'a göre, 1934
Rekabet modelinin (9.2) dezavantajları vardır; özellikle, iki türün bir arada yaşamasının ancak sayıları farklı faktörlerle sınırlıysa mümkün olduğu sonucu çıkar, ancak model, uzun vadeli bir arada yaşamayı sağlamak için farklılıkların ne kadar büyük olması gerektiğini göstermez. . Aynı zamanda değişen ortamda uzun süreli bir arada yaşayabilmek için belli bir büyüklüğe ulaşan bir farkın gerekli olduğu da bilinmektedir. Modele stokastik unsurların dahil edilmesi (örneğin, bir kaynak kullanım fonksiyonunun dahil edilmesi), bu sorunları niceliksel olarak araştırmamıza olanak tanır.
PREDATOR+VICTIM sistemi
 (9.7)
(9.7)
Burada (9.2)'den farklı olarak işaretler B 12 Ve B 21 farklıdır. Rekabet durumunda olduğu gibi, menşei
![]() (9.8)
(9.8)
kararsız düğüm tipinin özel bir noktasıdır. Diğer üç olası kararlı durum:
![]() ,(9.9)
,(9.9)
![]() (9.10)
(9.10)
![]() (9.11)
(9.11)
Böylece yalnızca avın hayatta kalması (9.10), yalnızca avcının (9.9) (başka besin kaynakları varsa) ve her iki türün bir arada yaşaması (9.11) mümkündür. Son seçeneği Ders 5'te zaten tartışmıştık. Bir avcı-av sistemi için olası faz portreleri Şekil 2'de sunulmaktadır. 9.4.
Yatay teğetlerin izoklinleri düz çizgilerdir
X 2 = – B 21 X 1 /C 2 + A 1/c2, X 2 = 0,
ve dikey teğetlerin izoklinleri- dümdüz
X 2 = – C 1 X 1 /B 12 + A 2 /B 12 , X 1 = 0.
Sabit noktalar dikey ve yatay teğet izoklinlerin kesişiminde bulunur.

Şek. 9.4 aşağıdakiler görülmektedir. Yırtıcı-av sistemi (9.7) istikrarlı bir denge pozisyonuna sahip olabilir;Ö Kurbanların Rum nüfusu tamamen tükendi ( ) ve yalnızca yırtıcılar kaldı (dönemŞekil 2'de 2. 9.4 A). Açıkçası, böyle bir durum ancak, söz konusu mağdurların türüne ek olarak, X 1 yırtıcı X 2 – ek güç kaynaklarına sahiptir. Bu gerçek modelde denklemin sağ tarafında x2 için pozitif terim ile yansıtılmaktadır. Özel noktalar(1) ve (3) (Şek. 9.4) A) kararsızdır. İkinci olasılık – Yırtıcı hayvan popülasyonunun tamamen tükendiği ve geriye yalnızca avın kaldığı istikrarlı ve durağan bir durum – kararlı nokta(3) (Şek. 9.4 6 ). Burada özel bir nokta var (1) – aynı zamanda kararsız bir düğüm.
Son olarak üçüncü olasılık – Yırtıcı ve av popülasyonlarının sürdürülebilir bir arada yaşaması (Şekil 1). 9.4 V), durağan sayıları formüllerle ifade edilenler (9.11).
Tek popülasyon durumunda olduğu gibi (bkz. Ders 3), model için (9.7) Stokastik bir model geliştirmek mümkündür ancak açık bir şekilde çözülemez. Bu nedenle kendimizi genel hususlarla sınırlayacağız. Örneğin denge noktasının eksenlerin her birinden belirli bir uzaklıkta bulunduğunu varsayalım. Daha sonra değerlerin üzerinde olduğu faz yörüngeleri içinX 1 , X 2 yeterince büyük kalırsa deterministik bir model oldukça tatmin edici olacaktır. Ancak faz yörüngesindeki bir noktada herhangi bir değişken çok büyük değilse, o zaman rastgele dalgalanmalar önemli hale gelebilir. Temsil eden noktanın eksenlerden birine hareket etmesine yol açarlar, bu da ilgili türlerin yok olması anlamına gelir.
Dolayısıyla stokastik modelin kararsız olduğu ortaya çıkıyor, çünkü stokastik "sürüklenme" er ya da geç türlerden birinin yok olmasına yol açıyor. Bu tür bir modelde, ya şans eseri ya da önce av popülasyonu ortadan kaldırıldığı için yırtıcı hayvanın nesli eninde sonunda tükenir. Yırtıcı-av sisteminin stokastik modeli Gause'un deneylerini iyi açıklıyor (Gause, 1934), hangi siliatlarda Paramettum kandatum başka bir siliat için av görevi gördü Didinium nasatum – yırtıcı hayvan. Deterministik denklemlere göre beklenen (9.7) bu deneylerdeki denge sayıları her türden yaklaşık olarak yalnızca beş bireydi; bu nedenle, tekrarlanan her deneyde avcıların veya avların (ve onlardan sonra avcıların) oldukça hızlı bir şekilde ölmesi şaşırtıcı değildir. Deneylerin sonuçları sunulmaktadır. incirde. 9.5.

Pirinç. 9.5. Yükseklik Parametyum kaudatum ve yırtıcı siliatlar Dadinium nasutum. İtibaren : Gause G.F. Varoluş mücadelesi. Baltimore, 1934
Dolayısıyla, Volterra'nın tür etkileşimi modellerinin analizi, bu tür sistemlerin davranış türlerinin çok çeşitli olmasına rağmen, rakip türlerin modelinde sayılardaki sönümsüz dalgalanmaların hiçbir şekilde olamayacağını göstermektedir. Ancak bu tür salınımlar doğada ve deneylerde gözlemlenmektedir. Teorik açıklamalara duyulan ihtiyaç, model açıklamalarının daha genel bir biçimde formüle edilmesinin nedenlerinden biriydi.
İki türün genelleştirilmiş etkileşim modelleri
Türlerin etkileşimini tanımlamak için çok sayıda model önerilmiştir; denklemlerin sağ tarafları, etkileşim halindeki popülasyonların sayısının fonksiyonlarıdır. Sabit dalgalanmalar da dahil olmak üzere geçici nüfus büyüklüğünün davranışını ne tür işlevlerin tanımlayabileceğini belirlemek için genel kriterlerin geliştirilmesi sorunu çözüldü. Bu modellerin en ünlüleri Kolmogorov (1935, gözden geçirilmiş makale - 1972) ve Rosenzweig'e (1963) aittir.
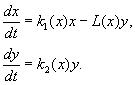 (9.12)
(9.12)
Model aşağıdaki varsayımları içerir:
1) Yırtıcı hayvanlar birbirleriyle etkileşime girmez; Yırtıcı üreme oranı k 2 ve kurban sayısı L birim zamanda bir yırtıcı tarafından yok edilmesi bağımlı değildir y.
2) Yırtıcı hayvanların varlığında av sayısındaki artış, yırtıcı hayvanların yokluğundaki artıştan yırtıcı hayvanlar tarafından yok edilen avların sayısının çıkarılmasıyla elde edilen sayıya eşittir. Fonksiyonlar k 1 (X), k 2 (X), L(X), süreklidir ve pozitif yarı eksende tanımlanır X, sen³ 0.
3) dk 1 /dx< 0. Bu, yırtıcı bir hayvanın yokluğunda avın üreme oranının, av sayısındaki artışla monoton bir şekilde azaldığı anlamına gelir; bu, yiyecek ve diğer kaynakların sınırlı mevcudiyetini yansıtır.
4) dk 2 /dx> 0, k 2 (0) < 0 < k 2 (¥ ). Av sayısının artmasıyla birlikte yırtıcı hayvanların üreme katsayısı, av sayısının artmasıyla birlikte monoton bir şekilde azalır. negatif değerler, (yiyecek hiçbir şey olmadığında) pozitife.
5) Bir avcı tarafından birim zamanda yok edilen av sayısı L(X)> 0 en N> 0; L(0)=0.
Sistemin (9.12) olası faz portreleri Şekil 2'de gösterilmektedir. 9.6:

Pirinç. 9.6.İki türün etkileşimini tanımlayan Kolmogorov sisteminin (9.12) faz portreleri farklı oranlar parametreler. Metindeki açıklamalar.
Sabit çözümler (iki veya üç tane vardır) aşağıdaki koordinatlara sahiptir:
(1). ` x=0;` y=0.
Herhangi bir parametre değeri için koordinatların kökeni bir eyerdir (Şekil 9.6 a-d).
(2). ` x=A,` y=0.(9.13)
Adenklemden belirlenir:
k 1 (A)=0.
Sabit çözüm (9.13) bir eyerdir, eğer B< A (Şekil 9.6 A, B, G), B denklemden belirlenir
k 2 (B)=0
(9.13) noktası pozitif çeyreğe yerleştirilir, eğer B>A . Bu kararlı bir düğüm .
Yırtıcı hayvanın ölümüne ve avın hayatta kalmasına karşılık gelen son durum Şekil 1'de gösterilmektedir. 9.6 V.
(3). ` x=B,` y=C.(9.14)
C'nin değeri denklemlerden belirlenir:
Nokta (9.14) – odaklama (Şek.9.6) A) veya düğüm (Şek.9.6) G), kararlılığı miktarın işaretine bağlıdırS
S 2 = – k 1 (B) –k 1 (B)B+L(B)C.
Eğer S>0, eğer bir nokta sabitseS<0 ‑ точка неустойчива, и вокруг нее могут существовать предельные циклы (рис. 9.6 B)
Yabancı literatürde Rosenzweig ve MacArthur (1963) tarafından önerilen benzer bir model daha sıklıkla dikkate alınmaktadır:
 (9.15)
(9.15)
Nerede F(X) - mağdur sayısındaki değişim oranı X yırtıcı hayvanların yokluğunda, F( x,y) - yırtıcılığın yoğunluğu, k- av biyokütlesinin yırtıcı biyokütleye dönüştürülmesinin verimliliğini karakterize eden katsayı, e- yırtıcı hayvan ölümleri.
Model (9.15), aşağıdaki varsayımlar altında Kolmogorov modelinin (9.12) özel bir durumuna indirgenir:
1) Yırtıcı hayvanların sayısı yalnızca av sayısıyla sınırlıdır,
2) belirli bir yırtıcı hayvanın avını yeme hızı yalnızca av popülasyonunun yoğunluğuna bağlıdır ve yırtıcı hayvan popülasyonunun yoğunluğuna bağlı değildir.
Daha sonra denklemler (9.15) formunu alır.

Gerçek türlerin etkileşimi açıklanırken denklemlerin sağ tarafları biyolojik gerçekliklerle ilgili fikirler doğrultusunda belirtilir. Bu türün en popüler modellerinden birini ele alalım.
İki tür böcek arasındaki etkileşim modeli (MacArthur, 1971)
Aşağıda ele alacağımız model, türlerden birinin erkeklerini kısırlaştırarak zararlı böcekleri kontrol etme konusundaki pratik sorunu çözmek için kullanıldı. Tür etkileşiminin biyolojik özelliklerine dayanarak aşağıdaki model yazılmıştır.
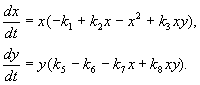 (9.16)
(9.16)
Burada x,y- iki tür böceğin biyokütlesi. Bu modelde tanımlanan türlerin trofik etkileşimleri çok karmaşıktır. Bu, denklemlerin sağ tarafındaki polinomların biçimini belirler.
İlk denklemin sağ tarafına bakalım. Böcek türleri X türün larvalarını yiyin en(üye +k 3 y), ama türün yetişkinleri en türün larvalarını yiyin X yüksek tür bolluğuna tabi X veya en veya her iki tür (üyeler –k 4 xy, – y 2). küçük X tür ölümü X doğal artışından daha yüksek (1 –k 1 +k 2 x–x 2 < 0 küçük X).İkinci denklemdeki terim k 5 Türün doğal gelişimini yansıtır y; –k 6 y – bu tür kendini kısıtlama,–k 7 X– türün larvalarını yemek en böcek türleri x, k 8 xy – tür biyokütlesindeki artış en türün yetişkin böcekleri tarafından tüketilmesi nedeniyle en türün larvaları X.
İncirde. 9.7 (9.16).

sistemin kararlı bir periyodik çözümünün yörüngesi olan bir limit döngüsü sunulur
Yönetim, popülasyon değerlerinde kısa vadeli, ani bir değişime indirgenebilir X Ve sen. Bu yöntem, bir veya her iki popülasyonun kimyasal yollarla tek seferlik yok edilmesi gibi kontrol yöntemlerine karşılık gelir. Yukarıda formüle edilen ifadeden, uyumlu popülasyonlar için bu kontrol yönteminin etkisiz olacağı açıktır, çünkü zamanla sistem yeniden durağan bir rejime ulaşacaktır.
Başka bir yol, örneğin sistem parametrelerinin değerlerini değiştirirken, görünümler arasındaki etkileşim fonksiyonlarının türünü değiştirmektir. Biyolojik kontrol yöntemlerinin karşılık geldiği parametrik yöntemdir. Böylece kısırlaştırılmış erkekler tanıtıldığında doğal nüfus artış hızı azalır. Aynı zamanda, yalnızca sıfır haşere sayısına sahip kararlı bir durağan durumun olduğu farklı türde bir faz portresi elde edersek, kontrol istenen sonuca yol açacaktır. – Zararlı bir türün popülasyonunun yok edilmesi. Bazen etkinin zararlının kendisine değil ortağına uygulanmasının tavsiye edilebileceğini belirtmek ilginçtir. Genel olarak hangi yöntemin daha etkili olduğunu söylemek imkansızdır. Bu, mevcut kontrollere ve popülasyonların etkileşimini tanımlayan fonksiyonların açık biçimine bağlıdır.
AD Bazykin'in modeli
Tür etkileşimi modellerinin teorik analizi en kapsamlı şekilde A.D. Bazykin’in “Etkileşimdeki Popülasyonların Biyofiziği” kitabında gerçekleştirildi (M., Nauka, 1985).
Bu kitapta incelenen yırtıcı-av modellerinden birini ele alalım.
 (9.17)
(9.17)
Sistem (9.17), yırtıcı doygunluğunun etkisini hesaba katan en basit Volterra avcı-av modelinin (5.17) bir genellemesidir. Model (5.17), av yoğunluğunun artmasıyla av otlatma yoğunluğunun doğrusal olarak arttığını varsaymaktadır; bu durum, yüksek av yoğunluklarında gerçeğe karşılık gelmemektedir. Bir yırtıcı hayvanın diyetinin av yoğunluğuna bağımlılığını tanımlamak için farklı işlevler seçilebilir. Seçilen fonksiyonun büyüme ile birlikte olması çok önemlidir. X asimptotik olarak sabit bir değere yöneldi. Model (9.6) lojistik bağımlılığı kullanmıştır. Bazykin modelinde hiperbol böyle bir fonksiyon olarak seçilmiştir X/(1+px). Bunun, mikroorganizmaların büyüme hızının substrat konsantrasyonuna bağımlılığını tanımlayan Monod formülünün formu olduğunu hatırlayalım. Burada av, substratın rolünü oynar ve avcı, mikroorganizmaların rolünü oynar. .
Sistem (9.17) yedi parametreye bağlıdır. Değişkenlerin değiştirilmesiyle parametre sayısı azaltılabilir:
X® (A/D)X; sen ® (A/D)/y;
T® (1 A)T; g (9.18)
ve dört parametreye bağlıdır.
Tam bir nitel çalışma için, dört boyutlu parametre uzayını farklı dinamik davranış türlerine sahip alanlara bölmek gerekir; Sistemin parametrik veya yapısal bir portresini oluşturun.
Daha sonra parametrik portrenin her alanı için faz portreleri oluşturmak ve parametrik portrenin farklı alanlarının sınırlarında faz portreleriyle meydana gelen çatallanmaları tanımlamak gerekir.
Tam bir parametrik portrenin oluşturulması, bazı parametrelerin sabit değerlerine sahip, düşük boyutlu bir parametrik portrenin bir dizi "dilimleri" (projeksiyonları) şeklinde gerçekleştirilir.
Sabit için sistemin parametrik portresi (9.18) G ve küçük eŞekil 9.8'de sunulmuştur. Portre, faz yörüngelerinin farklı davranış türlerine sahip 10 alan içermektedir.

Pirinç. 9.8.Sabit için sistemin parametrik portresi (9.18)G
ve küçük e
Sistemin farklı parametre oranlarındaki davranışı önemli ölçüde farklı olabilir (Şekil 9.9). Sistem şunları sağlar:
1) bir kararlı denge (bölge 1 ve 5);
2) bir kararlı limit çevrimi (bölge 3 ve 8);
3) iki kararlı denge (bölge 2)
4) kararlı limit döngüsü ve içindeki kararsız denge (bölgeler 6, 7, 9, 10)
5) kararlı limit döngüsü ve bunun dışında kararlı denge (bölge 4).
7, 9, 10 numaralı parametrik bölgelerde, dengenin çekim bölgesi, kararlı bir döngünün içinde yer alan kararsız bir sınır döngüsüyle sınırlıdır. En ilginç yapı, parametrik portrede 6. alana karşılık gelen faz portresidir. Şekil 2'de ayrıntılı olarak gösterilmektedir. 9.10.

Denge B2'nin (gölgeli) çekim alanı, kararsız odaktan B1 gelen bir "salyangoz" bükülmesidir. Başlangıçta sistemin B 1 civarında olduğu biliniyorsa, o zaman karşılık gelen yörüngenin B 2 dengesine mi yoksa üç denge noktası C'yi (eyer) çevreleyen kararlı bir sınır döngüsüne mi ulaşacağına karar vermek mümkündür. ), B 1 ve B 2 olasılıksal değerlendirmelere dayanmaktadır.
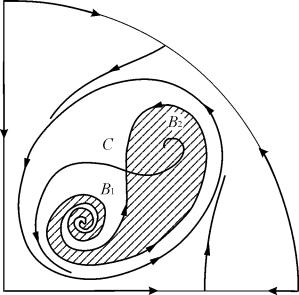
Şekil 9.10.Parametrik bölge 6 için sistem 9.18'in faz portresi. Çekim bölgesi B 2 gölgelidir
Parametrik bir portrede(9.7) 22 tane var oluşturan çeşitli çatallanma sınırları 7 çeşitli çatallanma türleri. Çalışmaları, parametreleri değiştiğinde olası sistem davranışı türlerini tanımlamamıza olanak tanır. Örneğin bölgeden taşınırken 1'den 3. bölgeye küçük bir limit döngüsünün doğuşu veya tek bir denge etrafında kendi kendine salınımların yumuşak doğuşu meydana gelir İÇİNDE. Kendi kendine salınımların benzer yumuşak doğuşu, ancak dengelerden biri etrafında, yani B 1 , bölgelerin sınırlarını geçerken ortaya çıkar 2 ve 4. Bölgeden ayrılırken 4'ten 5. bölgeye bir nokta etrafında kararlı limit döngüsüB 1 ayrılıkların döngüsünde “patlamalar” olur ve tek çekim noktası dengede kalır B 2 vesaire.
Uygulama açısından özellikle ilgi çekici olan, elbette, bir sistemin çatallanma sınırlarına yakınlığı için kriterlerin geliştirilmesidir. Aslında biyologlar doğal ekolojik sistemlerin “tamponlama” veya “esneklik” özelliğinin çok iyi farkındalar. Bu terimler genellikle bir sistemin dış etkileri absorbe etme yeteneğini ifade eder. Dış etkinin şiddeti belirli bir kritik değeri aşmadığı sürece sistemin davranışı niteliksel bir değişikliğe uğramaz. Faz düzleminde bu, sistemin kararlı bir denge durumuna veya parametreleri orijinalinden pek farklı olmayan kararlı bir sınır döngüsüne geri dönmesine karşılık gelir. Etkinin yoğunluğu izin verilen seviyeyi aştığında, sistem "bozulur" ve niteliksel olarak farklı bir dinamik davranış moduna geçer, örneğin basitçe yok olur. Bu fenomen bir çatallanma geçişine karşılık gelir.
Her çatallanma geçişi türünün, ekosistem için böyle bir geçişin tehlikesini değerlendirmeyi mümkün kılan kendine özgü özellikleri vardır. Tehlikeli bir sınırın yakınlığını gösteren bazı genel kriterler şunlardır: Tek tür örneğinde olduğu gibi, türlerden birinin sayısı azaldığında sistem, sayının çok yavaş bir şekilde başlangıç değerine dönmesiyle ifade edilen kararsız bir eyer noktasının yakınında "takılıp kalırsa", o zaman Sistem kritik sınıra yakındır. Av ve yırtıcı hayvan sayısındaki dalgalanmaların şeklindeki değişiklik de tehlikenin bir göstergesidir. Harmoniklere yakın salınımlar gevşeme haline gelirse ve salınımların genliği artarsa, bu durum sistemin stabilitesinin kaybolmasına ve türlerden birinin yok olmasına neden olabilir.
Türler arasındaki etkileşime ilişkin matematiksel teorinin daha da derinleştirilmesi, popülasyonların yapısının detaylandırılması ve zamansal ve mekansal faktörlerin dikkate alınması yönünde ilerlemektedir.
Edebiyat.
Kolmogorov A.N. Nüfus dinamiğinin matematiksel modellerinin nitel çalışması. // Sibernetiğin sorunları. M., 1972, Sayı 5.
MacArtur R. Ekolojik sistemlerin grafiksel analizi // Perinceton Üniversitesi biyoloji raporu bölümü. 1971
A.D. Bazykin “Etkileşen popülasyonların biyofiziği.” M., Nauka, 1985.
V. Volterra: “Varoluş mücadelesinin matematiksel teorisi.” M.. Bilim, 1976
Gause G.F. Varoluş mücadelesi. Baltimore, 1934.



