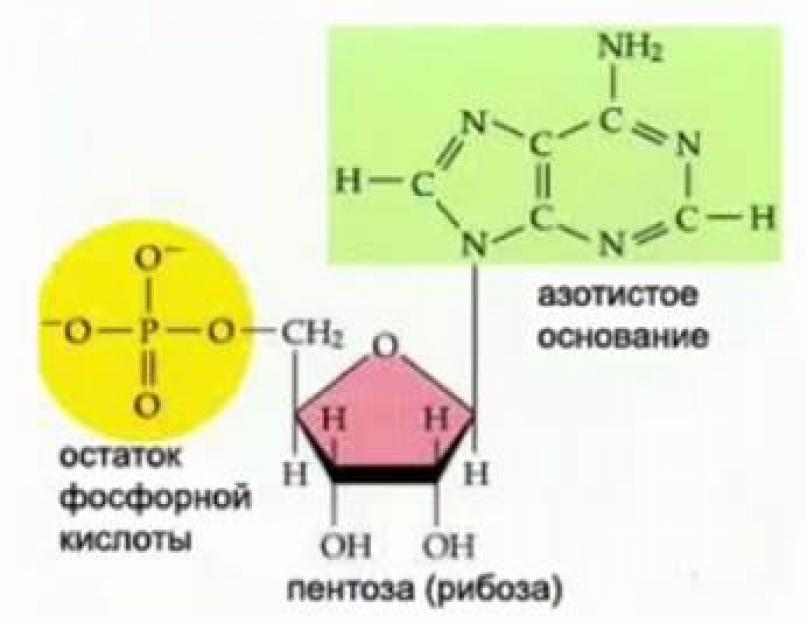
Мономеры РНК в составе нуклеотидов содержат пятиуглеродный сахар (пентоза), фосфорную кислоту (остаток фосфорной кислоты) и азотистое основание (см. Рис. 2).

Рис. 2. Строение нуклеотида РНК
Азотистые основания РНК - урацил, цитозин, аденин и гуанин. Моносахарид нуклеотида РНК представлен рибозой (см. Рис. 2).
РНК - одноцепочная молекула значительно меньших размеров, чем молекула ДНК.
Молекула РНК содержит от 75 до 10 000 нуклеотидов.

Рис. 3. РНК-содержащий вирус
Многие вирусы, например вирус гриппа, содержат в качестве единственной нуклеиновой кислоты молекулу РНК (см. Рис. 3). РНК-содержащих вирусов, болезнетворных для человека, больше, чем ДНК-содержащих. Они вызывают полиомиелит, гепатит А, острые простудные заболевания.
Арбовирусы - вирусы, которые переносятся членистоногими. Являются возбудителями клещевого и японского энцефалита, а также желтой лихорадки.
Реовирусы (см. Рис. 4), редкие возбудители респираторных и кишечных заболеваний человека, стали предметом особого научного интереса из-за того, что их генетический материал представлен в виде двухцепочной молекулы РНК.

Рис. 4. Строение реовируса
Также существуют ретровирусы, которые вызывают ряд онкологических заболеваний.
В зависимости от строения и выполняемой функции различают три основных типа РНК: рибосомную, транспортную и информационную (матричную).
1. Информационная РНК
Как показали исследования, информационная РНК составляет 3-5 % от общего содержания РНК в клетке. Это одноцепочная молекула, которая образовывается в процессе транскрипции на одной из цепей молекулы ДНК. Это связано с тем, что ДНК у ядерных организмов находятся в ядре, а синтез белка происходит на рибосомах в цитоплазме, поэтому возникла необходимость в «посреднике». Функцию «посредника» выполняет матричная РНК, она передает информацию о структуре белка из ядра клеток, где находится ДНК, к рибосомам, где эта информация реализуется (см. Рис. 5).

Рис. 5. Матричная РНК (мРНК)
В зависимости от объема копируемой информации, молекула матричной РНК может иметь различную длину.
Большинство матричных РНК существуют в клетке непродолжительное время. В бактериальных клетках существование таких РНК определяется минутами, а в клетках млекопитающих (в эритроцитах) синтез гемоглобина (белка) продолжается после утраты эритроцитами ядра в течение нескольких дней.
2. Рибосомная РНК
Рибосомные РНК (см. Рис. 6) составляют 80 % от всех рибосом, присутствующих в клетке. Эти РНК синтезируются в ядрышке, а в клетке они находятся в цитоплазме, где вместе с белками образуют рибосомы. На рибосомах происходит синтез белка. Здесь «код», заключенный в матричную РНК, транслируется в аминокислотную последовательность молекулы белка.

Рис. 6. Рибосомная РНК (рРНК)
3. Транспортная РНК
Транспортные РНК (см. Рис. 7) образуются в ядре на ДНК, а затем переходят в цитоплазму.

Рис. 7. Транспортная РНК (тРНК)
На долю таких РНК приходится около 10 % от общего содержания РНК в клетке. Они имеют самые короткие молекулы из 80-100 нуклеотидов.
Транспортные РНК присоединяют к себе аминокислоту и транспортируют ее к месту синтеза белка, к рибосомам.
Все известные транспортные РНК за счет комплементарного взаимодействия между азотистыми основаниями образовывают вторичную структуру, по форме напоминающую лист клевера (см. Рис. 8). В молекуле тРНК есть два активных участка - триплет антикодон на одном конце и акцепторный участок, присоединяющий аминокислоту, на другом.

Рис. 8. Строение тРНК («клеверный лист»)
Каждой аминокислоте соответствует комбинация из трех нуклеотидов, которая носит название триплет .

Рис. 9. Таблица генетического кода
Кодирующие аминокислоты триплеты - кодоны ДНК (см. Рис. 9) - передаются в виде информации триплетов (кодонов) мРНК. У верхушки клеверного листа тРНК располагается триплет нуклеотидов, который комплементарен соответствующему кодону мРНК (см. Рис. 10). Этот триплет различен для тРНК, переносящих разные аминокислоты, и кодирует именно ту аминокислоту, которая переносятся данной тРНК. Он получил название антикодон .

Рис. 10. тРНК
Акцепторный конец является «посадочной площадкой» для определенной аминокислоты.
Таким образом, различные типы РНК представляют собой единую функциональную систему, направленную на реализацию наследственной информации через синтез белка.
Концепция РНК мира заключается в том, что когда-то очень давно молекула РНК могла выполнять функцию как молекулы ДНК, так и белков.
В живых организмах практически все процессы происходят благодаря ферментам белковой природы. Белки, однако, не могут самореплицироваться и синтезируются в клетки на основании информации, заложенной в ДНК. Но и удвоение ДНК происходит только благодаря участию белков и РНК. Следовательно, образуется замкнутый круг, из-за которого в рамках теории возникновения жизни спонтанное возникновение такой сложной системы маловероятно.
В начале 1980-х годов в лаборатории ученых Чека и Олтмена (обладатели нобелевской премии по химии) в США была открыта каталитическая способность РНК. РНК-катализаторы были названы рибозимами (см. Рис. 11).

Рис. 11. Структура рибозимомолекулы РНК, выполняющей функцию катализа
Оказалось, что активный центр рибосом тоже содержит большое количество рибосомных РНК. Также РНК способны создавать двойную цепочку и самореплицироваться. То есть РНК могли существовать полностью автономно, катализируя метаболические реакции, например синтеза новых рибонуклеатидов, и самовоспроизводясь, сохраняя из поколения в поколение каталитические свойства. Накопление случайных мутаций привело к появлению РНК, катализирующих синтез определенных белков, являющихся более эффективными катализаторами, в связи с чем эти мутации закреплялись в ходе естественного отбора. Также возникли специализированные хранилища генетической информации - молекула ДНК, а РНК стала посредником между ДНК и белками.
Список литературы
- Каменский А.А., Криксунов Е.А., Пасечник В.В. Общая биология 10-11 класс Дрофа, 2005.
- Биология. 10 класс. Общая биология. Базовый уровень / П.В. Ижевский, О.А. Корнилова, Т.Е. Лощилина и др. - 2-е изд., переработанное. - Вентана-Граф, 2010. - 224 стр.
- Беляев Д.К. Биология 10-11 класс. Общая биология. Базовый уровень. - 11-е изд., стереотип. - М.: Просвещение, 2012. - 304 с.
- Агафонова И.Б., Захарова Е.Т., Сивоглазов В.И. Биология 10-11 класс. Общая биология. Базовый уровень. - 6-е изд., доп. - Дрофа, 2010. - 384 с.
- Orgchem.ru ().
- Appteka.ru ().
- Youtube.com ().
Домашнее задание
- Вопросы 4, 5 в конце параграфа 12 (стр. 52) - Каменский А.А., Криксунов Е.А., Пасечник В.В. «Общая биология», 10-11 класс ()
- Где в клетке находятся нуклеиновые кислоты?
Рибонуклеиновая кислота представляет собой сополимер пуриновых и пиримидиновых рибонуклеотидов, соединенных друг с другом, как и в ДНК, -фосфодиэфирными мостиками (рис. 37.6). Хотя эти два вида нуклеиновых кислот имеют много общего, по ряду признаков они отличаются друг от друга.
1. У РНК углеводным остатком, к которому присоединены пуриновые или пиримидиновые основания и фосфатные группы, является рибоза, а не 2-дезоксирибоза (как у ДНК).
2. Пиримидиновые компоненты РНК отличаются от таковых у ДНК. В состав РНК, как и в состав ДНК, входят нуклеотиды аденина, гуанина и цитозина. В то же время РНК (за исключением некоторых специальных случаев, на которых мы остановимся ниже) не содержит тимина, его место в молекуле РНК занимает урацил.
3. РНК - одноцепочечная молекула (в отличие от ДНК, имеющей двухцепочечную структуру), однако при наличии в цепи РНК участков с комплементарной последовательностью (противоположной полярности) единичная цепь РНК способна сворачиваться с образованием так называемых «шпилек», структур, имеющих двухспиральные характеристики (рис. 37.7).

Рис. 37.6. Фрагмент молекулы рибонуклеиновой кислоты (РНК), в котором пуриновые и пиримидиновые основания- аденин (А), урацил (U), цитозин (С) и гуанин (-удерживаются фосфодиэфирным остовом, соединяющим рибозильные остатки, связанные N-гликозидной связью с соответствующими нуклеиновыми основаниями. Обратите внимание: цепь РНК обладает определенной направленностью, на которую указывают 5- и З-концевые фосфатные остатки.
4. Так как молекула РНК представляет собой одиночную цепь, комплементарную только одной из цепей ДНК, содержание в ней гуанина не обязательно равно содержанию цитозина, а содержание аденина не обязательно равно содержанию урацила.
5. РНК может быть гидролизована щелочью до 2, З-циклических диэфиров мононуклеотидов; в роли промежуточного продукта гидролиза выступает 2, У, 5-триэфир, который не образуется при щелочном гидролизе ДНК из-за отсутствия у последней 2-гидроксильных групп; щелочная лабильность РНК (сравнительно с ДНК) является полезным свойством как для диагностических, так и для аналитических целей.
Информация, содержащаяся в одноцепочечной РНК, реализуется в виде определенной последовательности пуриновых и пиримидиновых оснований (т. е. в первичной структуре) полимерной цепи. Эта последовательность комплементарна кодирующей цепи гена, с которой «считывается» РНК. Вследствие комплементарности молекула РНК способна специфически связываться (гибридизоваться) с кодирующей цепью, но не гибридизуется с некодирующей цепью ДНК. Последовательность РНК (за исключением замены Т на U) идентична последовательности некодирующей цепи гена (рис. 37.8).
Биологические функции РНК
Известно несколько видов РНК. Почти все они непосредственно вовлечены в процесс биосинтеза белка. Молекулы цитоплазматической РНК, выполняющие функции матриц белкового синтеза, называются матричными РНК (мРНК). Другой вид цитоплазматической РНК-рибосомная РНК (рРНК) - выполняет роль структурных компонентов рибосом (органелл, играющих важную роль в синтезе белка). Адапторные молекулы транспортных РНК (тРНК) участвуют в трансляции (переводе) информации мРНК в последовательность аминокислот в белках.
Значительная часть РНК-первичных транскриптов, образующихся в эукариотических клетках, включая и клетки млекопитающих, - подвергается деградации в ядре и не играет какой-либо структурной или информационной роли в цитоплазме. В культивируемых

Рис. 37.7. Вторичная структура молекулы РНК типа «петли со стеблем» («шпилька»), возникающая вследствие внутримолекулярного образования водородных связей между комплементарными парами нуклеиновых оснований.
клетках человека обнаружен класс малых ядерных РНК которые непосредственно не участвуют в синтезе белка, но могут оказывать влияние на процессинг РНК и общую «архитектуру» клетки. Размеры этих относительно небольших молекул варьируют, последние содержат от 90 до 300 нуклеотидов (табл. 37.3).
РНК является основным генетическим материалом у некоторых вирусов животных и растений. Некоторые РНК-содержащие вирусы никогда не проходят стадию обратной транскрипции РНК в ДНК. Однако для большинства известных вирусов животных, таких, как ретровирусы, характерна обратная транскрипция их РНК-генома, направляемая РНК-зависимой ДНК-полимеразой (обратной транскриптазой) с образованием двухспиральной ДНК-копии. Во многих случаях образующийся двухспиральный ДНК-транскрипт встраивается в геном и в дальнейшем обеспечивает экспрессию генов вируса, а также наработку новых копий вирусных РНК-геномов.
Структурная организация РНК
Во всех эукариотических и прокариотических организмах существуют три основных класса молекул РНК: информационная (матричная или мессенджер) РНК (мРНК), транспортная (тРНК) и рибосомная (рРНК). Представители этих классов отличаются друг от друга размерами, функциями и стабильностью.
Информационная (мРНК) - наиболее гетерогенный в отношении размеров и стабильности класс. Все представители этого класса служат переносчиками информации от гена к белок-синтезирующей системе клетки. Они выполняют роль матриц для синтезируемого полипептида, т. е. определяют аминокислотную последовательность белка (рис. 37.9).
Информационные РНК, особенно эукариотические, обладают некоторыми уникальными структурными особенностями. 5-Конец мРНК «кэпирован» 7-метилгуанозинтрифосфатом, присоединенным к 5-гидроксилу соседнего 2-0-метилрибонуклеозида через остаток трифосфата (рис. 37.10). Молекулы мРНК часто содержат внутренние остатки 6-метиладенина и 2-0-метилированные рибонуклеотиды. Хотя смысл «кэпирования» до конца еще не выяснен, можно предположить, что образующаяся структура 5-конца мРНК используется для специфического узнавания в системе трансляции. Синтез белка начинается на 5"-(кэпированном) конце мРНК. Другой конец большинства молекул мРНК (З-конец) содержит полиаденилатную цепочку из 20-250 нуклеотидов. Специфические функции этого окончательно не установлены. Можно предполагать, что данная структура отвечает за поддержание внутриклеточной стабильности мРНК. Некоторые мРНК, включая гистоновые не содержат poly (А). Наличие poly (А) в структуре мРНК используется для отделения от других видов РНК посредством фракционирования тотальной РНК на колонках с oligo (Т), иммобилизованным на твердом носителе типа целлюлозы. Связывание мРНК с колонкой происходит за счет комплементарных взаимодействий poly (А)-«хвоста» с иммобилизованным oligo (Т).
Рис. 37.8. Последовательность гена и его РНК-транскрипта. Показаны кодирующая и некодирующая цепи, и отмечена их полярность. РНК-транскрипт, имеющий полярность комплементарен кодирующей цепи (с полярностью 3 - 5) и идентичен по последовательности (за исключением замен Т на U) и полярности некодирующей цепи ДНК.

Рис. 37.9. Экспрессия генетической информации ДНК в форме мРНК-транскрипта и последующая трансляция при участии рибосом с образованием специфической молекулы белка.
(см. скан)
Рис. 37.10. Структура «кэпа», находящегося на 5-конце большинства эукариотических матричных РНК 7-метилгуанозинтрифосфат присоединяется к 5-концу мРНК. на котором обычно находится 2-О-метилпуриновый нуклеотид.
В клетках млекопитающих, включая клетки человека, зрелые молекулы мРНК, находящиеся в цитоплазме, не являются полной копией транскрибируемого участка гена. Образующийся в результате транскрипции полирибонуклеотид представляет собой предшественник цитоплазматической мРНК, перед выходом из ядра он подвергается специфическому процессингу. Непроцессированные продукты транскрипции, обнаруживаемые в ядрах клеток млекопитающих, образуют четвертый класс молекул РНК. Такие ядерные РНК очень гетерогенны и достигают значительных размеров. Молекулы гетерогенных ядерных РНК могут иметь молекулярную массу более , в то время как молекулярная масса мРНК обычно не превышает 2106. подвергаются процессингу в ядре, и образующиеся зрелые мРНК поступают в цитоплазму, где служат матрицей для биосинтеза белка.
Молекулы транспортных РНК (тРНК) обычно содержат около 75 нуклеотидов. Молекулярная масса таких молекул составляет . тРНК также формируются в результате специфического процессинга соответствующих молекул-предшественников (см. гл. 39). Транспортные тРНК выполняют функцию посредников в ходе трансляции мРНК. В любой клетке присутствуют не менее 20 видов молекул тРНК. Каждый вид (иногда несколько видов) тРНК соответствует одной из 20 аминокислот, необходимых для синтеза белка. Хотя каждая специфическая тРНК отличается от других нуклеотидной последовательностью, все они имеют и общие черты. Благодаря нескольим внутрицепочечным комплементарным участкам, все тРНК обладают вторичной структурой, получившей название «клеверный лист» (рис. 37.11).
Молекулы всех видов тРНК имеют четыре основных плеча. Акцепторное плечо состоит из «стебля» спаренных нуклеотидов и заканчивается последовательностью ССА Именно через У-гидроксильную группу аденозильного остатка происходит связывание с карбоксильной группой аминокислоты. Остальные плечи тоже состоят из «стеблей», образованных комплементарными парами оснований, и петель из неспаренных оснований (рис. 37.7). Антикодоновое плечо узнает нуклеотидный триплет или кодон (см. гл. 40) в мРНК. D-плечо названо так из-за наличия в нем дигидроуридина, -плечо названо по последовательности Т-псевдоуридин-С. Дополнительное плечо представляет собой наиболее вариабельную структуру и служит основой классификации тРНК. тРНК класса 1 (75% от общего их числа) обладают дополнительным плечом длиной 3-5 пар оснований. Дополнительное плечо у тРНК-молекул класса 2 состоит из 13-21 пар оснований и часто включает неспаренную петлю.

Рис. 37.11. Структура молекулы аминоацил-тРНК, к 3-ССА-концу которой присоединена аминокислота . Указаны внутримолекулярные водородные связи и расположение антикодонового, ТТС- и дигидроурацилового плеч. (From J. D. Watson. Molecular biology of the Gene 3rd, ed.. Copyright 1976, 1970, 1965 by W. A. Benjamin, Inc., Menlo Park Calif.)
Вторичная структура, определяемая системой комплементарных взаимодействий нуклеотидных оснований соответствующих плеч, характерна для всех видов Акцепторное плечо содержит семь пар оснований, -плечо - пять пар оснований, плечо D - три (или четыре) пары оснований.
Молекулы тРНК весьма стабильны у прокариот и несколько менее стабильны у эукариот. Обратная ситуация характерна для мРНК, которая довольно нестабильна у прокариот, а у эукариотических организмов обладает значительной стабильностью.
Рибосомная РНК. Рибосома - это цитоплазматическая нуклеопротеиновая структура, предназначенная для синтеза белка по мРНК-матрице. Рибосома обеспечивает специфический контакт в результате которого и происходит трансляция нуклеотидной последовательности, считанной с определенного гена, в аминокислотную последовательность соответствующего белка.
В табл. 37.2 представлены компоненты рибосом млекопитающих, имеющих молекулярную массу 4,210 6 и скорость седиментации (единиц Сведберга). Рибосомы млекопитающих состоят из двух нуклеопротеиновых субъединиц - большой с
Таблица 37.2. Компоненты рибосом млекопитающих

молекулярной массой (60S), и малой, имеющей молекулярную массу (40S). 608-субъединица содержит 58-рибосомную РНК (рРНК), 5,8S-pPHK и 28S-pPHK, а также более 50 различных полипептидов. Малая, 408-субъединица включает единственную 18S-pPHK и около 30 полипептидных цепей. Все рибосомные РНК, за исключением 5S-PHK, имеют общего предшественника-45S-PHK, локализованную в ядрышке (см. гл. 40). У молекулы 5S-PHK предшественник собственный. В ядрышке происходит упаковка высокометилированных рибосомных РНК с рибосомными белками. В цитоплазме рибосомы достаточно устойчивы и способны осуществлять большое число циклов трансляции.
Небольшие стабильные РНК. В эукариотических клетках обнаружено большое число дискретных, высококонсервативных, небольших и стабильных молекул РНК. Большинство РНК этого типа обнаруживаются в составе рибонуклеопротеинов и локализованы в ядре, цитоплазме или одновременно в обоих компартментах. Размеры этих молекул варьируют от 90 до 300 нуклеотидов, содержание их - 100000-1000000 копий на клетку.
Малые ядерные нуклеопротеиновые частицы (часто называемые snurps - от англ. small nuclear ribonucleic particles), вероятно, играют существенную роль в регуляции экспрессии генов. Нуклеопротеиновые частицы типа U7, по-видимому, участвуют в формировании З-концов гистоновых мРНК. Частицы , вероятно, необходимы для полиаденилирования, a - для удаления интронов и процессинга мРНК (см. гл. 39). Табл. 37.3. суммирует некоторые характеристики небольших стабильных РНК.
Таблица 37.3. Некоторые виды небольших стабильных РНК, обнаруженные в клетках млекопитающих

ЛИТЕРАТУРА
Darnell J. et al. Molecular Cell Biology, Scientific American Books, 1986.
Hunt T. DNA Makes RNA Makes Protein, Elsevier, 1983. Lewin B. Genes, 2nd ed., Wiley, 1985.
Rich A. et al. The chemistry and biology of left-handed Z-DNA, Annu. Rev. Biochem., 1984, 53, 847.
Turner P. Controlling roles for snurps, Nature, 1985, 316, 105. Watson J. D. The Double Helix, Atheneum, 1968.
Watson J. D., Crick F.H.C. Molecular structure of nucleic acids. Nature, 1953, 171, 737.
Zieve G. W. Two groups of small stable RNAs, Cell, 1981, 25, 296.
Кандидат биологических наук С. ГРИГОРОВИЧ.
На самой ранней заре своей истории, когда человек приобрел разум, а с ним и способность к абстрактному мышлению, он стал пленником непреодолимой потребности все объяснить. Почему светят Солнце и Луна? Почему текут реки? Как устроен мир? Безусловно, одним из самых главных был вопрос о сути живого. Резкое отличие живого, растущего, от мертвого, неподвижного, слишком бросалось в глаза, чтобы его можно было проигнорировать.
Первый вирус, описанный Д. Ивановским в 1892 году, - вирус табачной мозаики. Благодаря этому открытию стало ясно, что существуют живые создания более примитивные, чем клетка.
Русский микробиолог Д. И. Ивановский (1864-1920), основоположник вирусологии.
В 1924 году А. И. Опарин (1894-1980) высказал предположение, что в атмосфере молодой Земли, состоявшей из водорода, метана, аммиака, углекислого газа и паров воды, могли синтезироваться аминокислоты, которые затем спонтанно соединились в белки.
Американский биолог Освальд Эвери убедительно продемонстрировал в опытах с бактериями, что именно нуклеиновые кислоты отвечают за передачу наследственных свойств.
Сравнительная структура РНК и ДНК.
Двумерная пространственная структура рибозима простейшего организма Tetrahymena.
Схематическое изображение рибосомы - молекулярной машины для синтеза белка.
Схема процесса "эволюции в пробирке" (селекс-метод).
Луи Пастер (1822-1895) первым обнаружил, что кристаллы одного и того же вещества - винной кислоты - могут иметь две зеркально-симметричные пространственные конфигурации.
В начале 1950-х годов Стенли Миллер из Чикагского университета (США) проделал первый эксперимент, моделирующий химические реакции, которые могли протекать в условиях молодой Земли.
Хиральные молекулы, например аминокислоты, зеркально симметричны, как левая и правая рука. Сам термин "хиральность" происходит от греческого слова "хирос" - рука.
Теория РНК-мира.
Наука и жизнь // Иллюстрации
На каждом этапе истории люди предлагали свое решение загадки появления жизни на нашей планете. Древние, не знавшие слова "наука", находили для неизвестного простое и доступное объяснение: "Все, что есть вокруг, было когда-то и кем-то создано". Так появились боги.
Со времен зарождения древних цивилизаций в Египте, Китае, затем и в колыбели современной науки - Греции, вплоть до Средних веков, основным методом познания мира служили наблюдения и мнения "авторитетов". Постоянные наблюдения однозначно свидетельствовали, что живое при соблюдении определенных условий появляется из неживого: комары и крокодилы - из болотной тины, мухи - из гниющей пищи, а мыши - из грязного белья, пересыпанного пшеницей. Важно лишь соблюсти определенную температуру и влажность.

Европейские "ученые" Средневековья, опираясь на религиозную догму о сотворении мира и непостижимости божественных замыслов, считали возможным спорить о зарождении жизни только в рамках Библии и религиозных писаний. Суть сотворенного Богом невозможно постичь, а можно лишь "уточнить", пользуясь сведениями из священных текстов или находясь под влиянием божественного вдохновения. Проверять гипотезы в то время считалось плохим тоном, и всякая попытка подвергнуть сомнению мнение святой церкви рассматривалась как дело небогоугодное, ересь и святотатство.
Познание жизни топталось на месте. Вершиной научной мысли в течение двух тысяч лет оставались достижения философов Древней Греции. Наиболее значимыми из них были Платон (428/427 - 347 гг. до н. э.) и его ученик Аристотель (384 - 322 гг. до н. э.). Платон в числе прочего предложил идею одушевления изначально неживой материи благодаря вселению в нее бессмертной нематериальной души - "психеи". Так появилась теория самозарождения живого из неживого.
Великое для науки слово "эксперимент" пришло с эпохой Возрождения. Две тысячи лет понадобились для того, чтобы человек решился усомниться в непреложности авторитетных утверждений ученых древности. Одним из первых смельчаков, известных нам, стал итальянский врач Франсиско Реди (1626 - 1698). Он провел чрезвычайно простой, но эффектный опыт: поместив в несколько сосудов по куску мяса, одни из них накрыл плотной тканью, другие - марлей, а третьи оставил открытыми. Тот факт, что личинки мух развивались только в открытых сосудах (на которые могли садиться мухи), но не в закрытых (к которым все же был доступ воздуха), резко противоречил верованиям сторонников Платона и Аристотеля о непостижимой жизненной силе, носящейся в воздухе и превращающей неживую материю в живую.
Этот и подобные ему опыты положили начало периоду ожесточенных сражений между двумя группами ученых: виталистами и механистами. Суть спора состояла в вопросе: "Может ли функционирование (и появление) живого быть объяснено физическими законами, применимы ми также и к неживой материи?" Виталисты отвечали на него отрицательно. "Клетка - только из клетки, все живое - только от живого!" Это положение, выдвинутое в середине XIX века, стало знаменем витализма. Самое парадоксальное в этом споре то, что даже сегодня, зная о "неживой" природе составляющих наш организм атомов и молекул и в общем согласившись с механистической точкой зрения, ученые не имеют экспериментального подтверждения возможности зарождения клеточной жизни из неодушевленной материи. Никому еще не удалось "составить" даже самую примитивную клетку из "неорганических", присутствующих вне живых организмов, "деталей". А значит, окончательную точку в этом эпохальном споре еще предстоит поставить.
Так как же могла возникнуть жизнь на Земле? Разделяя позиции механистов, легче всего конечно же представить, что жизнь сначала должна была возникнуть в какой-нибудь очень простой, примитивно устроенной форме. Но, несмотря на простоту строения, это все же должна быть Жизнь, то есть то, что обладает минимальным набором свойств, отличающих живое от неживого.
Каковы же они, эти критически важные для жизни свойства? Что, собственно, отличает живое от неживого?
До конца XIX века ученые были убеждены, что все живое построено из клеток, и это является самым очевидным отличием его от неживой материи. Так считали до открытия вирусов, которые, хотя и меньше всех известных клеток, могут активно заражать другие организмы, размножаться в них и производить потомство, обладающее такими же (или очень похожими) биологическими свойствами. Первый из обнаруженных вирусов, вирус табачной мозаики, описан русским ученым Дмитрием Ивановским (1864-1920) в 1892 году. С той поры стало ясно, что более примитивные создания, чем клетки, могут также претендовать на право называться Жизнью.
Открытие вирусов, а затем и еще более примитивных форм живого - вироидов позволило в итоге сформулировать минимальный набор свойств, которые необходимы и достаточны, чтобы исследуемый объект можно было назвать живым. Во-первых, он должен быть способен к воспроизводству себе подобных. Это, однако, не единственное условие. Если бы гипотетическая первородная субстанция жизни (например, примитивная клетка или молекула) была способна лишь просто производить свои точные копии, она в итоге не смогла бы выжить в меняющихся условиях окружающей среды на молодой Земле и образование других, более сложных форм (эволюция) стало бы невозможным. Следовательно, нашу предполагаемую примитивную "субстанцию первожизни" можно определить как нечто, устроенное максимально просто, но при этом способное изменяться и передавать свои свойства потомкам.
РНК состоит, как правило, из одной цепи, закрученной в спираль. У вирусов есть двухцепочные РНК. РНК содержится в ядрышке, ядре, цитоплазме, рибосомах. Молекулы РНК менее короткие, чем молекулы ДНК.
Типы РНК
Различают три типа РНК: рибосомную, матричную (информационную – иРНК), транспортную (тРНК). Они отличаются между собою местонахождением в клетке, размерами, нуклеотидным составом и функциональными свойствами.
Синтезируется РНК с участием ферментов РНК-полимераз на молекуле ДНК. Последовательность нуклеотидов участка молекулы ДНК определяет порядок расположения нуклеотидов в молекуле РНК.
У большинства клеток содержание РНК значительно большее (от 5 до 10 раз), чем содержание ДНК. Наибольшая часть РНК приходится на рибосомную.
Функции РНК
Функции РНК : реализует наследственную информацию, принимает участие в синтезе белков.
Информационная (матричная) РНК (иРНК) представляет собой копию участка ДНК, то есть одного или нескольких генов. Она переносит генетическую информацию к месту синтеза полипептидной цепи и принимает в нем непосредственное участие. Соответственно длине участка ДНК, которое и РНК копирует, она состоит из 300-30 000 нуклеотидов. Часть и РНК в клетке составляет около 5 % общего количества. Молекулы и РНК относительно нестабильные – быстро распадаются на нуклеотиды. Срок их жизни составляет в клетках эукариот до нескольких часов, у микроорганизмов – несколько минут.
Подобно молекуле ДНК, и РНК имеет вторичную и третичную структуры, которые формируются с помощью водородных связей, гидрофобных, электростатических взаимодействий и т. п.
Рибосомная РНК составляет 60 % массы рибосом, около 85 % общего количества РНК клетки. Включает 3000-5000 нуклеотидов. Она не принимает участия в передаче наследственной информации. Входит в состав рибосомы и взаимодействует с ее белками, которых у эукариот около 100. У эукариот есть четыре типа рибосомной РНК, у прокариот - три. Выполняет структурную функцию: обеспечивает определенное пространственное расположение иРНК и тРНК на рибосоме.
Транспортная (тРНК) - переносит аминокислоты к месту синтеза белка. По принципу комплементарности узнает участок иРНК, отвечающий аминокислоте, которая транспортируется. К месту синтеза белка каждая аминокислота транспортируется своей тРНК. тРНК транспортируются элементами цитоскелета клетки.
Имеет форму трехлистника (листка клевера) - постоянную вторичную структуру, которая обеспечивается водородными связями. На верхушке тРНК расположен триплет нуклеотидов, соответствующий кодону иРНК и называющийся антикодоном . Возле основания есть участок, к которому благодаря ковалентной связи молекула аминокислоты прикрепляется. Содержит тРНК 70-90 нуклеотидов. Составляет до 10 % общего количества РНК. Известно около 60 видов тРНК.
тРНК может иметь довольно компактную L-подобной неправильной формы третичную структуру.
Динуклеотиды
Состоят из двух нуклеотидов, но имеют особенности в строении. Наиболее известными являются: никотинамидадениндинуклеотид (НАД +), никотинамидадениндинуклеотидфосфат (НАДФ +). Главной функцией является перенесение электронов (2) и иона водорода (1). Могут восстанавливаться:
НАД + + 2е — + Н + →НАДН;
НАДФ + + 2е — + Н + →НАДФН.
На определенном участке для некоторых реакций эти соединения отдают протон водорода, электроны:
НАДН →НАД + + 2е — + Н + ;
НАДФН → НАДФ + + 2е — + Н +
В зависимости от того, какой моносахарид содержится в структурном звене полинуклеотида - рибоза или 2-дезоксирибоза , различают
- рибонуклеиновые кислоты (РНК) и
- дезоксирибонуклеиновые кислоты (ДНК).
Нуклеотидные звенья макромолекул ДНК могут содержать аденин, гуанин, цитозин и тимин . Состав РНК отличается тем, что вместо тимина присутствует урацил .
Молекулярная масса ДНК достигает десятков миллионов а.е.м. Это самые длинные из известных макромолекул. Значительно меньше молекулярная масса РНК (от нескольких сотен до десятков тысяч). ДНК содержатся в основном в ядрах клеток, РНК в рибосомах и протоплазме клеток.
При описании строения нуклеиновых кислот учитывают различные уровни организации макромолекул: первичную и вторичную структуру.
- Первичная структура нуклеиновых кислот это нуклеотидный состав и определенная последовательность нуклеотидных звеньев в полимерной цепи.
В сокращённом однобуквенном обозначении эта структура записывается как ...– А – Г – Ц –...
- Под вторичной структурой нуклеиновых кислот понимают пространственно упорядоченные формы полинуклеотидных цепей.

Такая пространственная структура удерживается множеством водородных связей, образуемых азотистыми основаниями, направленными внутрь спирали. Водородные связи возникают между пуриновым основанием одной цепи и пиримидиновым основанием другой цепи. Эти основания составляют комплементарные пары (от лат. complementum - дополнение). Образование водородных связей между комплементарными парами оснований обусловлено их пространственным соответствием. Пиримидиновое основание комплементарно пуриновому основанию:

Водородные связи между другими парами оснований не позволяют им разместиться в структуре двойной спирали. Таким образом,
- ТИМИН (Т) комплементарен АДЕНИНУ (А),
- ЦИТОЗИН (Ц) комплементарен ГУАНИНУ (Г).

Комплементарность полинуклеотидных цепей служит химической основой главной функции ДНК хранения и передачи наследственных признаков.
Способность ДНК не только хранить, но и использовать генетическую информацию определяется следующими ее свойствами:
Вторичная структура РНК . В отличие от ДНК, молекулы РНК состоят из одной полинуклеотидной цепи и не имеют строго определенной пространственной формы (вторичная структура РНК зависит от их биологических функций).
Основная роль РНК непосредственное участие в биосинтезе белка. Известны три вида клеточных РНК, которые отличаются по местоположению в клетке, составу, размерам и свойствам, определяющим их специфическую роль в образовании белковых макромолекул:
- информационные (матричные) РНК передают закодированную в ДНК информацию о структуре белка от ядра клетки к рибосомам, где и осуществляется синтез белка;
- транспортные РНК собирают аминокислоты в цитоплазме клетки и переносят их в рибосому; молекулы РНК этого типа "узнают" по соответствующим участкам цепи информационной РНК, какие аминокислоты должны участвовать в синтезе белка;
- рибосомные РНК обеспечивают синтез белка определенного строения, считывая информацию с информационной (матричной) РНК.



