الكود الجيني هو نظام لتسجيل المعلومات الوراثية في جزيئات الحمض النووي ، بناءً على تناوب معين لتسلسلات النيوكليوتيدات في الحمض النووي أو الحمض النووي الريبي التي تشكل أكواد تتوافق مع الأحماض الأمينية في البروتين.
خصائص الكود الجيني.
يحتوي الكود الجيني على عدة خصائص.
الثلاثية.
الانحطاط أو التكرار.
غموض.
قطبية.
غير التداخل.
الاكتناز.
براعه.
وتجدر الإشارة إلى أن بعض المؤلفين يقدمون أيضًا خصائص أخرى للشفرة تتعلق بالسمات الكيميائية للنيوكليوتيدات المدرجة في الكود أو بتكرار حدوث الأحماض الأمينية الفردية في بروتينات الجسم ، إلخ. ومع ذلك ، فإن هذه الخصائص تتبع ما سبق ، لذلك سننظر فيها هناك.
أ. الثلاثية. يحتوي الكود الجيني ، مثل العديد من الأنظمة المنظمة بشكل معقد ، على أصغر وحدة هيكلية وأصغر وحدة وظيفية. الثلاثي هو أصغر وحدة هيكلية للشفرة الجينية. يتكون من ثلاثة نيوكليوتيدات. الكودون هو أصغر وحدة وظيفية في الشفرة الجينية. كقاعدة عامة ، تسمى ثلاثة توائم مرنا الكودونات. في الشفرة الجينية ، يؤدي الكودون عدة وظائف. أولاً ، وظيفته الرئيسية هي أنه يرمز لحمض أميني واحد. ثانيًا ، قد لا يرمز الكودون لحمض أميني ، ولكن في هذه الحالة له وظيفة مختلفة (انظر أدناه). كما يتضح من التعريف ، الثلاثي هو المفهوم الذي يميز ابتدائي الوحدة الهيكليةالكود الجيني (ثلاثة نيوكليوتيدات). الكودون يميز الوحدة الدلالية الابتدائيةالجينوم - تحدد ثلاثة نيوكليوتيدات الارتباط بسلسلة عديد الببتيد لحمض أميني واحد.
تم فك شفرة الوحدة البنائية الأولية نظريًا ، ثم تم تأكيد وجودها تجريبيًا. في الواقع ، لا يمكن تشفير 20 من الأحماض الأمينية بواحد أو اثنين من النيوكليوتيدات. الأخيرة هي فقط 4. ثلاثة من كل أربعة نيوكليوتيدات تعطي 4 3 = 64 متغيرًا ، وهو ما يغطي أكثر من عدد الأحماض الأمينية الموجودة في الكائنات الحية (انظر الجدول 1).
مجموعات النيوكليوتيدات الواردة في الجدول 64 لها ميزتان. أولاً ، من بين 64 نوعًا مختلفًا من ثلاثة توائم ، 61 فقط عبارة عن كودونات وترميز أي حمض أميني ، يطلق عليهم أكواد المعنى. ثلاثة توائم لا يتم ترميزها
الأحماض الأمينية هي إشارات توقف تحدد نهاية الترجمة. هناك ثلاثة توائم من هذا القبيل UAA ، UAG ، UGA، يطلق عليهم أيضًا اسم "لا معنى له" (أكواد لا معنى لها). نتيجة لطفرة مرتبطة باستبدال أحد النوكليوتيدات في ثلاثي بأخرى ، يمكن أن ينشأ كودون لا معنى له من كودون حاسي. يسمى هذا النوع من الطفرات طفرة هراء. إذا تم تشكيل إشارة التوقف هذه داخل الجين (في الجزء المعلوماتي منه) ، فسيتم إيقاف العملية باستمرار أثناء تخليق البروتين في هذا المكان - سيتم فقط تصنيع الجزء الأول (قبل إشارة التوقف) من البروتين. سيواجه الشخص المصاب بمثل هذا المرض نقصًا في البروتين وسيواجه الأعراض المرتبطة بهذا النقص. على سبيل المثال ، تم العثور على هذا النوع من الطفرات في الجين المشفر لسلسلة بيتا الهيموجلوبين. يتم تصنيع سلسلة هيموجلوبين مختصرة غير نشطة ، والتي يتم تدميرها بسرعة. نتيجة لذلك ، يتم تكوين جزيء الهيموجلوبين الخالي من سلسلة بيتا. من الواضح أن مثل هذا الجزيء من غير المرجح أن يؤدي واجباته بالكامل. هناك مرض خطير يتطور حسب نوع فقر الدم الانحلالي (بيتا-صفر ثلاسيميا ، مشتق من الكلمة اليونانية "تالاس" - البحر الأبيض المتوسط ، حيث تم اكتشاف هذا المرض لأول مرة).
تختلف آلية عمل أكواد الإيقاف عن آلية عمل أكواد المعنى. هذا يأتي من حقيقة أنه بالنسبة لجميع الكودونات التي تشفر الأحماض الأمينية ، تم العثور على الحمض الريبي النووي النقال المقابل. لم يتم العثور على tRNAs لشفرات هراء. لذلك ، لا يشارك الحمض الريبي النووي النقال في عملية وقف تخليق البروتين.
كودونأغسطس (أحيانًا GUG في البكتيريا) لا يشفر فقط الأحماض الأمينية ميثيونين وفالين ، ولكن أيضًابادئ البث .
ب. الانحطاط أو التكرار.
61 من 64 رمزًا ثلاثيًا لـ 20 حمضًا أمينيًا. تشير هذه الزيادة بمقدار ثلاثة أضعاف في عدد التوائم الثلاثة على عدد الأحماض الأمينية إلى أنه يمكن استخدام خيارين من خيارات الترميز في نقل المعلومات. أولاً ، لا يمكن أن تشارك جميع الكودونات الـ 64 في ترميز 20 من الأحماض الأمينية ، ولكن 20 فقط من الأحماض الأمينية ، وثانيًا ، يمكن تشفير الأحماض الأمينية بعدة أكواد. أظهرت الدراسات أن الطبيعة تستخدم الخيار الأخير.
تفضيله واضح. إذا كان 20 فقط من أصل 64 متغيرًا ثلاثيًا متورطًا في ترميز الأحماض الأمينية ، فسيظل 44 ثلاثيًا (من أصل 64) غير مشفر ، أي لا معنى لها (أكواد هراء). في وقت سابق ، أشرنا إلى مدى خطورة تحول ثلاثي الترميز نتيجة للطفرة إلى كودون لا معنى له على حياة الخلية - وهذا يعطل بشكل كبير العملية الطبيعية لبوليميراز الحمض النووي الريبي ، مما يؤدي في النهاية إلى تطور الأمراض. يوجد حاليًا ثلاثة أكواد غير منطقية في جينومنا ، والآن تخيل ماذا سيحدث إذا زاد عدد الكودونات غير المنطقية بنحو 15 مرة. من الواضح أنه في مثل هذه الحالة ، سيكون انتقال الكودونات العادية إلى أكواد غير منطقية أعلى بما لا يقاس.
يُطلق على الكود الذي يتم فيه ترميز أحد الأحماض الأمينية بواسطة عدة ثلاثة توائم اسم متدهور أو فائض عن الحاجة. يحتوي كل حمض أميني تقريبًا على العديد من الكودونات. لذلك ، يمكن ترميز ليسين الأحماض الأمينية بستة توائم - UUA ، UUG ، CUU ، CUC ، CUA ، CUG. يتم ترميز الفالين بأربعة توائم ، فينيل ألانين باثنين وفقط التربتوفان والميثيونينمشفر بواسطة كودون واحد. يتم استدعاء الخاصية المرتبطة بتسجيل نفس المعلومات بأحرف مختلفة انحطاط.
يرتبط عدد الكودونات المخصصة لحمض أميني واحد ارتباطًا جيدًا بتكرار حدوث الحمض الأميني في البروتينات.
وهذا على الأرجح ليس عرضيًا. كلما زاد تواتر حدوث حمض أميني في البروتين ، كلما تم تمثيل كودون هذا الحمض الأميني في الجينوم ، كلما زادت احتمالية تلفه بواسطة عوامل مطفرة. لذلك ، من الواضح أن الكودون المتحول يكون أكثر احتمالا لتشفير نفس الحمض الأميني إذا كان شديد الانحلال. من هذه المواقف ، فإن انحلال الشفرة الجينية هو آلية تحمي الجينوم البشري من التلف.
وتجدر الإشارة إلى أن مصطلح الانحلال يستخدم في علم الوراثة الجزيئي بمعنى آخر أيضًا. نظرًا لأن الجزء الرئيسي من المعلومات في الكودون يقع على النيوكليوتيدات الأولين ، فقد تبين أن القاعدة في الموضع الثالث من الكودون قليلة الأهمية. هذه الظاهرة تسمى "انحلال القاعدة الثالثة". تقلل الميزة الأخيرة من تأثير الطفرات. على سبيل المثال ، من المعروف أن الوظيفة الرئيسية لخلايا الدم الحمراء هي نقل الأكسجين من الرئتين إلى الأنسجة وثاني أكسيد الكربون من الأنسجة إلى الرئتين. يتم تنفيذ هذه الوظيفة عن طريق الصباغ التنفسي - الهيموغلوبين ، الذي يملأ السيتوبلازم بأكمله في كريات الدم الحمراء. يتكون من جزء بروتيني - غلوبين ، يتم ترميزه بواسطة الجين المقابل. بالإضافة إلى البروتين ، يحتوي الهيموغلوبين على الهيم الذي يحتوي على الحديد. تؤدي الطفرات في جينات الغلوبين إلى ظهور أنواع مختلفة من الهيموجلوبين. في أغلب الأحيان ، ترتبط الطفرات بـ استبدال نيوكليوتيد بآخر وظهور كودون جديد في الجين، والتي يمكن أن ترمز إلى حمض أميني جديد في سلسلة الهيموجلوبين البولي ببتيد. في ثلاثة توائم ، نتيجة لطفرة ، يمكن استبدال أي نوكليوتيد - الأول أو الثاني أو الثالث. من المعروف أن عدة مئات من الطفرات تؤثر على سلامة جينات الغلوبين. قرب 400 منها مرتبطة باستبدال النيوكليوتيدات المفردة في الجين واستبدال الأحماض الأمينية المقابلة في عديد الببتيد. من هؤلاء فقط 100 تؤدي البدائل إلى عدم استقرار الهيموجلوبين وأنواع مختلفة من الأمراض من خفيفة إلى شديدة للغاية. 300 (حوالي 64٪) من الطفرات البديلة لا تؤثر على وظيفة الهيموغلوبين ولا تؤدي إلى أمراض. أحد أسباب ذلك هو "انحلال القاعدة الثالثة" المذكورة أعلاه ، عندما يؤدي استبدال النيوكليوتيد الثالث في الترميز الثلاثي السيرين ، والليوسين ، والبرولين ، والأرجينين ، وبعض الأحماض الأمينية الأخرى إلى ظهور كودون مرادف ترميز نفس الأحماض الأمينية. ظاهريًا ، لن تظهر مثل هذه الطفرة نفسها. في المقابل ، فإن أي استبدال للنيوكليوتيدات الأولى أو الثانية في ثلاثة توائم في 100٪ من الحالات يؤدي إلى ظهور متغير جديد للهيموجلوبين. ولكن حتى في هذه الحالة ، قد لا تكون هناك اضطرابات نمطية شديدة. والسبب في ذلك هو استبدال حمض أميني في الهيموجلوبين بحمض أميني آخر مشابه للأول من حيث الخصائص الفيزيائية والكيميائية. على سبيل المثال ، إذا تم استبدال حمض أميني بخصائص محبة للماء بحمض أميني آخر ، ولكن بنفس الخصائص.
يتكون الهيموغلوبين من مجموعة بورفيرين الحديد من الهيم (يرتبط بها جزيئات الأكسجين وثاني أكسيد الكربون) وبروتين - غلوبين. يحتوي الهيموغلوبين البالغ (HbA) على نوعين متطابقين - سلاسل واثنين -السلاسل. مركب - تحتوي السلسلة على 141 بقايا من الأحماض الأمينية ، - سلسلة - 146 ، - و - تختلف السلاسل في العديد من بقايا الأحماض الأمينية. يتم ترميز تسلسل الأحماض الأمينية لكل سلسلة غلوبين بواسطة الجين الخاص بها. ترميز الجينات - تقع السلسلة على الذراع القصيرة للكروموسوم 16 ، -جين - في الذراع القصيرة للكروموسوم 11. تغيير في ترميز الجينات - تؤدي سلسلة الهيموجلوبين من النوكليوتيدات الأولى أو الثانية دائمًا تقريبًا إلى ظهور أحماض أمينية جديدة في البروتين ، وتعطيل وظائف الهيموجلوبين وعواقب وخيمة على المريض. على سبيل المثال ، استبدال "C" في واحد من ثلاثة توائم CAU (الهيستدين) بـ "U" سيؤدي إلى ظهور UAU ثلاثي جديد يشفر حمض أميني آخر - التيروزين. ظاهريًا ، سيظهر هذا في مرض خطير .. أ استبدال مماثل في الموضع 63 - سلسلة من بولي ببتيد الهيستيدين إلى التيروزين ستزعزع استقرار الهيموغلوبين. يتطور مرض ميتهيموغلوبينية الدم. تغيير ، نتيجة للطفرة ، حمض الجلوتاميك إلى حمض الفالين في الموضع السادس السلسلة هي سبب مرض شديد - فقر الدم المنجلي. دعونا لا نكمل القائمة المحزنة. نلاحظ فقط أنه عند استبدال النيوكليوتيدات الأولين ، قد يظهر الحمض الأميني مشابهًا في الخواص الفيزيائية والكيميائية للحمض السابق. وهكذا ، فإن استبدال النيوكليوتيد الثاني في واحد من ثلاثة توائم يشفر حمض الجلوتاميك (GAA) في يؤدي -chain على "Y" إلى ظهور عنصر ترميز ثلاثي جديد (GUA) فالين ، ويشكل استبدال النوكليوتيدات الأول بـ "A" ثلاثي AAA يشفر الحمض الأميني ليسين. يتشابه حمض الجلوتاميك والليسين في الخصائص الفيزيائية والكيميائية - كلاهما محبة للماء. فالين حمض أميني كاره للماء. لذلك ، فإن استبدال حمض الغلوتاميك المحب للماء بفالين كاره للماء يغير بشكل كبير خصائص الهيموجلوبين ، مما يؤدي في النهاية إلى تطور فقر الدم المنجلي ، في حين أن استبدال حمض الغلوتاميك المحب للماء مع ليسين ماء يغير وظيفة الهيموغلوبين إلى حد أقل - المرضى تطوير شكل خفيف من فقر الدم. نتيجة لاستبدال القاعدة الثالثة ، يمكن للثلاثية الجديدة ترميز نفس الأحماض الأمينية مثل سابقتها. على سبيل المثال ، إذا تم استبدال اليوراسيل بالسيتوزين في CAC الثلاثي ونشأ ثلاثي CAC ، فلن يتم اكتشاف أي تغييرات نمطية في الشخص عمليًا. هذا أمر مفهوم لأن يرمز كلا التوائم الثلاثة لنفس الحمض الأميني ، الهيستيدين.
في الختام ، من المناسب التأكيد على أن انحطاط الكود الجيني وانحلال القاعدة الثالثة من الوضع البيولوجي العام هي آليات وقائية مدمجة في التطور في التركيب الفريد للحمض النووي الريبي (DNA) والحمض النووي الريبي (RNA).
في. غموض.
كل ثلاثة توائم (باستثناء تلك التي لا معنى لها) يشفر حمض أميني واحد فقط. وهكذا ، في اتجاه الكودون - الأحماض الأمينية ، فإن الكود الجيني لا لبس فيه ، في اتجاه الحمض الأميني - الكودون - غامض (متحلل).
خالية من الغموض
حمض أميني كودون
تتدهور
وفي هذه الحالة ، فإن الحاجة إلى عدم الغموض في الشفرة الجينية واضحة. في متغير آخر ، أثناء ترجمة نفس الكودون ، سيتم إدخال أحماض أمينية مختلفة في سلسلة البروتين ، ونتيجة لذلك ، سيتم تكوين بروتينات ذات هياكل أولية مختلفة ووظائف مختلفة. سيتحول التمثيل الغذائي للخلية إلى طريقة عمل "جين واحد - عدة عديدات ببتيدات". من الواضح أنه في مثل هذه الحالة ستفقد الوظيفة التنظيمية للجينات تمامًا.
ز - القطبية
قراءة المعلومات من DNA و mRNA تحدث في اتجاه واحد فقط. تعد القطبية ضرورية لتحديد الهياكل ذات الترتيب الأعلى (الثانوية ، والثالثية ، وما إلى ذلك). تحدثنا سابقًا عن حقيقة أن الهياكل ذات الترتيب الأدنى تحدد الهياكل ذات الترتيب الأعلى. تتشكل البنية والتراكيب الثلاثية ذات الترتيب الأعلى في البروتينات على الفور بمجرد أن تتحرك سلسلة RNA المركبة بعيدًا عن جزيء DNA أو تتحرك سلسلة polypeptide بعيدًا عن الريبوسوم. بينما يكتسب الطرف الحر من RNA أو polypeptide بنية ثلاثية ، لا يزال الطرف الآخر من السلسلة يتم تصنيعه على DNA (إذا تم نسخ RNA) أو الريبوسوم (إذا تم نسخ عديد الببتيد).
لذلك ، فإن العملية أحادية الاتجاه لقراءة المعلومات (في تخليق الحمض النووي الريبي والبروتين) ضرورية ليس فقط لتحديد تسلسل النيوكليوتيدات أو الأحماض الأمينية في المادة المركبة ، ولكن من أجل التحديد الصارم للثانوي والثالث وما إلى ذلك. الهياكل.
هـ - عدم التداخل.
قد يتداخل الرمز وقد لا يتداخل. في معظم الكائنات الحية ، تكون الشفرة غير متداخلة. تم العثور على رمز متداخل في بعض العاثيات.
جوهر الكود غير المتداخل هو أن نيوكليوتيد أحد الكودون لا يمكن أن يكون نيوكليوتيد كودون آخر في نفس الوقت. إذا كانت الشفرة متداخلة ، فإن تسلسل سبعة نيوكليوتيدات (GCUGCUG) لا يمكن أن يشفر اثنين من الأحماض الأمينية (ألانين ألانين) (الشكل 33 ، أ) كما في حالة الشفرة غير المتداخلة ، ولكن ثلاثة (إذا كان نيوكليوتيد واحد شائع) (الشكل 33 ، ب) أو خمسة (إذا كان هناك نيوكليوتيدان شائعان) (انظر الشكل 33 ، ج). في الحالتين الأخيرتين ، قد تؤدي طفرة أي نوكليوتيد إلى انتهاك في تسلسل حالتين ، ثلاثة ، إلخ. أحماض أمينية.
ومع ذلك ، فقد وجد أن طفرة في نوكليوتيد واحد تعطل دائمًا تضمين حمض أميني واحد في عديد الببتيد. هذه حجة مهمة لصالح حقيقة أن الكود غير متداخل.
دعونا نوضح هذا في الشكل 34. تظهر الخطوط العريضة ثلاثة توائم ترميز الأحماض الأمينية في حالة الشفرة غير المتداخلة والمتداخلة. أظهرت التجارب بشكل لا لبس فيه أن الشفرة الجينية غير متداخلة. دون الخوض في تفاصيل التجربة ، نلاحظ أنه إذا استبدلنا النيوكليوتيد الثالث في تسلسل النيوكليوتيدات (انظر الشكل 34)في (مميزة بعلامة النجمة) إلى البعض الآخر ثم:
1. مع رمز غير متداخل ، سيكون للبروتين الذي يتحكم فيه هذا التسلسل بديلاً عن حمض أميني واحد (أول) (مميز بعلامة النجمة).
2. مع وجود رمز متداخل في الخيار A ، سيحدث الاستبدال في اثنين (الأول والثاني) من الأحماض الأمينية (المميزة بعلامات النجمة). بموجب الخيار ب ، سيؤثر الاستبدال على ثلاثة أحماض أمينية (مميزة بعلامات النجمة).
ومع ذلك ، فقد أظهرت العديد من التجارب أنه عندما يتم كسر نوكليوتيد واحد في الحمض النووي ، فإن البروتين يؤثر دائمًا على حمض أميني واحد فقط ، وهو أمر نموذجي للشفرة غير المتداخلة.
ГЦУГЦУГ ГЦУГЦУГ ГЦУГЦУГ
HCC HCC HCC UHC CUG HCC CUG UGC HCC CUG
*** *** *** *** *** ***
Alanine - Alanine Ala - Cys - Lei Ala - Lei - Lei - Ala - Lei
أ ب ج
رمز غير متداخل متداخل
أرز. 34. مخطط يشرح وجود رمز غير متداخل في الجينوم (شرح في النص).
يرتبط عدم تداخل الشفرة الجينية بخاصية أخرى - تبدأ قراءة المعلومات من نقطة معينة - إشارة البدء. إشارة البدء هذه في mRNA هي ترميز الميثيونين AUG.
وتجدر الإشارة إلى أن الشخص لا يزال لديه عدد قليل من الجينات التي تنحرف عن القاعدة العامة وتتداخل.
ه. الاكتناز.
لا توجد علامات ترقيم بين الكودونات. بمعنى آخر ، لا يتم فصل الثلاثة توائم عن بعضها البعض ، على سبيل المثال ، بواسطة نوكليوتيد واحد لا معنى له. تم إثبات عدم وجود "علامات الترقيم" في الشفرة الجينية في التجارب.
و. براعه.
الرمز هو نفسه لجميع الكائنات الحية التي تعيش على الأرض. تم الحصول على دليل مباشر على عالمية الكود الجيني بمقارنة تسلسل الحمض النووي بتسلسلات البروتين المقابلة. اتضح أن نفس مجموعات قيم الكود تُستخدم في جميع الجينومات البكتيرية وحقيقية النواة. هناك استثناءات ، لكنها ليست كثيرة.
تم العثور على الاستثناءات الأولى لعالمية الكود الجيني في الميتوكوندريا لبعض الأنواع الحيوانية. يتعلق هذا بكودون الإنهاء UGA ، الذي يقرأ نفس كودون UGG الذي يشفر الحمض الأميني التربتوفان. كما تم العثور على انحرافات أخرى نادرة عن العالمية.
نظام كود الحمض النووي.
يتكون الكود الجيني للحمض النووي من 64 ثلاثة توائم من النيوكليوتيدات. هذه الثلاثة توائم تسمى الكودونات. يرمز كل كودون لواحد من الأحماض الأمينية العشرين المستخدمة في تخليق البروتين. هذا يعطي بعض التكرار في الكود: يتم ترميز معظم الأحماض الأمينية بأكثر من كودون واحد.
يؤدي أحد الكودون وظيفتين مترابطتين: فهو يشير إلى بداية الترجمة ويرمز إلى دمج ميثيونين الأحماض الأمينية (Met) في سلسلة البولي ببتيد المتنامية. تم تصميم نظام ترميز الحمض النووي بحيث يمكن التعبير عن الشفرة الجينية إما كرموز RNA أو كرموز DNA. تحدث أكواد الحمض النووي الريبي في الحمض النووي الريبي (مرنا) وهذه الكودونات قادرة على قراءة المعلومات أثناء تخليق بولي ببتيدات (عملية تسمى الترجمة). لكن كل جزيء mRNA يكتسب تسلسل نيوكليوتيد في النسخ من الجين المقابل.
يمكن ترميز جميع الأحماض الأمينية باستثناء اثنين (Met و Trp) من 2 إلى 6 أكواد مختلفة. ومع ذلك ، فإن جينوم معظم الكائنات الحية يظهر أن بعض الكودونات مفضلة على غيرها. في البشر ، على سبيل المثال ، يتم ترميز الألانين بواسطة دول مجلس التعاون الخليجي أربع مرات أكثر من تشفيره في GCG. ربما يشير هذا إلى كفاءة ترجمة أكبر لجهاز الترجمة (على سبيل المثال ، الريبوسوم) لبعض الكودونات.

الشيفرة الجينية عالمية تقريبا. يتم تخصيص نفس الكودونات لنفس امتداد الأحماض الأمينية ونفس إشارات البداية والتوقف هي نفسها إلى حد كبير في الحيوانات والنباتات والكائنات الحية الدقيقة. ومع ذلك ، تم العثور على بعض الاستثناءات. يتضمن معظمها تخصيص واحد أو اثنين من أكواد الإيقاف الثلاثة إلى حمض أميني.
وزارة التعليم والعلوم في الوكالة الفيدرالية الروسية للتعليم
المؤسسة التعليمية الحكومية للتعليم المهني العالي "جامعة ألتاي التقنية الحكومية تحمل اسم I.I. Polzunov"
قسم العلوم الطبيعية وتحليل النظام
مقال حول موضوع "الكود الجيني"
1. مفهوم الشيفرة الجينية
3. المعلومات الجينية
فهرس
1. مفهوم الشيفرة الجينية
الكود الجيني هو نظام موحد لتسجيل المعلومات الوراثية في جزيئات الحمض النووي في شكل سلسلة من النيوكليوتيدات ، مميزة للكائنات الحية. يُشار إلى كل نوكليوتيد بحرف كبير يبدأ باسم القاعدة النيتروجينية التي تشكل جزءًا منه: - أ (أ) الأدينين ؛ - G (G) جوانين ؛ - C (C) السيتوزين ؛ - T (T) الثايمين (في DNA) أو U (U) uracil (في mRNA).
يتم تنفيذ الكود الجيني في الخلية على مرحلتين: النسخ والترجمة.
أولها يحدث في النواة. يتكون من تخليق جزيئات الرنا المرسال في الأقسام المقابلة من الحمض النووي. في هذه الحالة ، يتم "إعادة كتابة" تسلسل نوكليوتيدات الحمض النووي في تسلسل نيوكليوتيدات الحمض النووي الريبي. المرحلة الثانية تحدث في السيتوبلازم ، على الريبوسومات. في هذه الحالة ، يتم ترجمة تسلسل النوكليوتيدات لـ i-RNA إلى تسلسل الأحماض الأمينية في البروتين: تستمر هذه المرحلة بمشاركة RNA (t-RNA) والإنزيمات المقابلة.
2. خصائص الكود الجيني
1. الثلاثية
يتم ترميز كل حمض أميني بواسطة سلسلة من 3 نيوكليوتيدات.
الثلاثي أو الكودون هو سلسلة من ثلاثة نيوكليوتيدات ترمز لحمض أميني واحد.
لا يمكن أن يكون الرمز أحاديًا ، لأن 4 (عدد النيوكليوتيدات المختلفة في الحمض النووي) أقل من 20. لا يمكن أن يكون الرمز مزدوجًا ، لأن 16 (عدد التوليفات والتباديلات لـ 4 نيوكليوتيدات في 2) أقل من 20. يمكن أن يكون الرمز ثلاثيًا ، لأن 64 (عدد التوليفات والتبديلات من 4 إلى 3) أكبر من 20.
2. الانحطاط.
يتم ترميز جميع الأحماض الأمينية باستثناء الميثيونين والتريبتوفان بأكثر من ثلاثة توائم: 2 أحماض أمينية 1 ثلاثية = 2 9 أحماض أمينية 2 ثلاثية لكل منها = 18 1 حمض أميني 3 توائم = 3 5 أحماض أمينية 4 ثلاثة توائم لكل منها = 20 3 أحماض أمينية 6 ثلاثة توائم كل منها = 18 إجمالي 61 رمزًا ثلاثيًا لـ 20 حمضًا أمينيًا.
3. وجود علامات الترقيم بين الجينات.
الجين هو جزء من الحمض النووي يرمز لسلسلة بولي ببتيد واحدة أو جزيء واحد من الحمض النووي الريبي ، أو الرنا الريباسي ، أو الرنا الريباسي.
جينات الرنا الريباسي و الرنا الريباسي و الرنا الرناوي لا ترمز للبروتينات.
في نهاية كل جين يقوم بتشفير بولي ببتيد ، يوجد واحد على الأقل من 3 أكواد إنهاء ، أو إشارات توقف: UAA ، UAG ، UGA. قاموا بإنهاء البث.
بشكل تقليدي ، ينتمي كودون AUG أيضًا إلى علامات الترقيم - الأولى بعد تسلسل القائد. ينفذ وظيفة الحرف الكبير. في هذا الموقف ، يتم ترميز الفورميل ميثيونين (في بدائيات النوى).
4. التفرد.
كل ثلاثة توائم يشفر حمض أميني واحد فقط أو عبارة عن فاصل ترجمة.
الاستثناء هو كودون AUG. في بدائيات النوى ، في الموضع الأول (حرف كبير) يرمز إلى فورميل ميثيونين ، وفي أي موضع آخر يرمز للميثيونين.
5. الضغط ، أو عدم وجود علامات الترقيم داخل الجين.
داخل الجين ، كل نوكليوتيد هو جزء من كودون مهم.
في عام 1961 أثبت سيمور بينزر وفرانسيس كريك تجريبياً أن الشفرة ثلاثية ومضغوطة.

جوهر التجربة: الطفرة "+" - إدخال نوكليوتيد واحد. "-" الطفرة - فقدان أحد النوكليوتيدات. طفرة واحدة "+" أو "-" في بداية الجين تفسد الجين بأكمله. كما أن الطفرة المزدوجة "+" أو "-" تفسد الجين بأكمله. الطفرة الثلاثية "+" أو "-" في بداية الجين تفسد جزءًا فقط منه. الطفرة الرباعية "+" أو "-" تفسد الجين بأكمله مرة أخرى.
أثبتت التجربة أن الشفرة ثلاثية ولا توجد علامات ترقيم داخل الجين. أجريت التجربة على جينين متجاورين من الملتهمة وأظهرت بالإضافة إلى ذلك وجود علامات ترقيم بين الجينات.
3. المعلومات الجينية
المعلومات الجينية هي برنامج لخصائص الكائن الحي ، يتم استلامها من أسلافها ومضمنة في هياكل وراثية في شكل شفرة جينية.
من المفترض أن تكون المعلومات الوراثية تتم وفقًا للمخطط: العمليات الجيوكيميائية - تكوين المعادن - التحفيز التطوري (التحفيز الذاتي).
من الممكن أن تكون الجينات البدائية الأولى عبارة عن بلورات جريزوفولفين من الطين ، وتصطف كل طبقة جديدة من الطين وفقًا للسمات الهيكلية السابقة ، كما لو كانت تتلقى معلومات حول التركيب منها.
يحدث إدراك المعلومات الوراثية في عملية تخليق جزيئات البروتين بمساعدة ثلاثة أنواع من الحمض النووي الريبي: المعلوماتية (مرنا) ، والنقل (الحمض الريبي النووي النقال) والريبوزوم (الرنا الريباسي). تمر عملية نقل المعلومات: - من خلال قناة الاتصال المباشر: DNA - RNA - بروتين ؛ و- عبر قناة التغذية الراجعة: البيئة - البروتين - الحمض النووي.
الكائنات الحية قادرة على تلقي المعلومات وتخزينها ونقلها. علاوة على ذلك ، تميل الكائنات الحية إلى استخدام المعلومات الواردة عن نفسها والعالم من حولها بأكبر قدر ممكن من الكفاءة. تنتقل المعلومات الوراثية المضمنة في الجينات والضرورية للكائن الحي للوجود والتطور والتكاثر من كل فرد إلى نسله. تحدد هذه المعلومات اتجاه تطور الكائن الحي ، وفي عملية تفاعله مع البيئة ، يمكن أن يتشوه رد الفعل تجاه فرده ، وبالتالي ضمان تطور تطور الأحفاد. في عملية تطور الكائن الحي ، تظهر معلومات جديدة ويتم تذكرها ، بما في ذلك قيمة المعلومات التي تزداد.
في سياق تنفيذ المعلومات الوراثية في ظل ظروف بيئية معينة ، يتم تشكيل النمط الظاهري للكائنات الحية لنوع بيولوجي معين.
تحدد المعلومات الجينية التركيب المورفولوجي ، والنمو ، والتطور ، والتمثيل الغذائي ، والمستودع العقلي ، والاستعداد للأمراض والعيوب الوراثية في الجسم.
لاحظ العديد من العلماء ، عن حق ، دور المعلومات في تكوين الكائنات الحية وتطورها ، أن هذا الظرف هو أحد المعايير الرئيسية للحياة. لذا ، ف. يعتقد كاراجودين أن: "الحياة هي مثل هذا الشكل من أشكال وجود المعلومات والهياكل المشفرة بها ، مما يضمن استنساخ هذه المعلومات في ظروف بيئية مناسبة". كما لاحظت أ. أ. ارتباط المعلومات بالحياة. ليابونوف: "الحياة هي حالة من المادة شديدة التنظيم تستخدم المعلومات المشفرة بواسطة حالات الجزيئات الفردية لتطوير تفاعلات مستمرة." قام عالم الفيزياء الفلكية المعروف ن. يؤكد كارداشيف أيضًا على المكون المعلوماتي للحياة: "تنشأ الحياة بسبب إمكانية تخليق نوع خاص من الجزيئات التي يمكن أن تتذكر وتستخدم في البداية أبسط المعلومات حول البيئة وبنيتها الخاصة ، والتي يستخدمونها للحفاظ على الذات. ، للتكاثر ، وهو أمر مهم بشكل خاص بالنسبة لنا للحصول على مزيد من المعلومات ". يلفت عالم البيئة ف. تيبلر الانتباه إلى قدرة الكائنات الحية على تخزين ونقل المعلومات في كتابه "فيزياء الخلود": "أنا أعرّف الحياة على أنها نوع من المعلومات المشفرة التي يحفظها الانتقاء الطبيعي". علاوة على ذلك ، يعتقد أنه إذا كان الأمر كذلك ، فإن نظام معلومات الحياة أبدي ولانهائي وخالد.
أظهر اكتشاف الشفرة الجينية ووضع قوانين البيولوجيا الجزيئية الحاجة إلى الجمع بين علم الوراثة الحديث ونظرية التطور الداروينية. وهكذا ، وُلد نموذج بيولوجي جديد - نظرية التطور التركيبية (STE) ، والتي يمكن اعتبارها بالفعل بيولوجيا غير كلاسيكية.
الأفكار الرئيسية لتطور داروين مع ثالوثه - الوراثة ، والتنوع ، والاختيار الطبيعي - في النظرة الحديثة لتطور العالم الحي تكملها أفكار ليس فقط عن الانتقاء الطبيعي ، ولكن من هذا الاختيار ، الذي يتم تحديده وراثيًا. يمكن اعتبار بداية تطور التطور التركيبي أو العام من عمل S. Chetverikov على علم الوراثة السكانية ، حيث تبين أنه لا تخضع السمات الفردية والأفراد للاختيار ، ولكن النمط الجيني لجميع السكان ، ولكن يتم تنفيذه من خلال السمات المظهرية للأفراد الأفراد. هذا يؤدي إلى انتشار التغييرات المفيدة بين السكان. وهكذا ، يتم تنفيذ آلية التطور من خلال الطفرات العشوائية على المستوى الجيني ، ومن خلال وراثة الصفات الأكثر قيمة (قيمة المعلومات!) ، والتي تحدد تكيف الصفات الطفرية مع البيئة ، وتوفير النسل الأكثر قابلية للحياة. .
تؤدي التغيرات المناخية الموسمية ، ومختلف الكوارث الطبيعية أو التي من صنع الإنسان ، من ناحية ، إلى تغيير في تواتر تكرار الجينات بين السكان ، ونتيجة لذلك ، إلى انخفاض في التباين الوراثي. تسمى هذه العملية أحيانًا بالانحراف الجيني. ومن ناحية أخرى ، إلى تغييرات في تركيز الطفرات المختلفة وانخفاض في تنوع الأنماط الجينية الموجودة في السكان ، مما قد يؤدي إلى تغييرات في اتجاه وشدة إجراء الانتقاء.
4. فك الشفرة الوراثية البشرية
في مايو 2006 ، نشر العلماء الذين يعملون على فك شفرة الجينوم البشري خريطة جينية كاملة للكروموسوم 1 ، والذي كان آخر كروموسوم بشري غير كامل التسلسل.
نُشرت خريطة جينية بشرية أولية في عام 2003 ، إيذانا بنهاية رسمية لمشروع الجينوم البشري. في إطاره ، تم تسلسل شظايا الجينوم التي تحتوي على 99 ٪ من الجينات البشرية. كانت دقة تحديد الجينات 99.99٪. ومع ذلك ، في نهاية المشروع ، تم تسلسل أربعة كروموسومات فقط من أصل 24 كروموسومًا بشكل كامل. الحقيقة هي أنه بالإضافة إلى الجينات ، تحتوي الكروموسومات على أجزاء لا تشفر أي سمات ولا تشارك في تخليق البروتين. لا يزال الدور الذي تلعبه هذه الشظايا في حياة الكائن الحي غير معروف ، لكن المزيد والمزيد من الباحثين يميلون إلى الاعتقاد بأن دراستهم تتطلب اهتمامًا وثيقًا.
المحاضرة 5 الكود الجيني
تعريف المفهوم
الكود الجيني هو نظام لتسجيل المعلومات حول تسلسل الأحماض الأمينية في البروتينات باستخدام تسلسل النيوكليوتيدات في الحمض النووي.
نظرًا لأن الحمض النووي لا يشارك بشكل مباشر في تخليق البروتين ، فإن الشفرة مكتوبة بلغة RNA. يحتوي الحمض النووي الريبي على اليوراسيل بدلاً من الثايمين.
خصائص الكود الجيني
1. الثلاثية
يتم ترميز كل حمض أميني بواسطة سلسلة من 3 نيوكليوتيدات.
التعريف: الثلاثي أو الكودون هو سلسلة من ثلاثة نيوكليوتيدات ترمز لحمض أميني واحد.
لا يمكن أن يكون الرمز أحاديًا ، لأن 4 (عدد النيوكليوتيدات المختلفة في الحمض النووي) أقل من 20. لا يمكن أن يكون الرمز مزدوجًا ، لأن 16 (عدد التوليفات والتباديلات لـ 4 نيوكليوتيدات في 2) أقل من 20. يمكن أن يكون الرمز ثلاثيًا ، لأن 64 (عدد التوليفات والتبديلات من 4 إلى 3) أكبر من 20.
2. الانحطاط.
يتم ترميز جميع الأحماض الأمينية ، باستثناء الميثيونين والتربتوفان ، بأكثر من ثلاثة توائم:
2 AKs لثلاثة توائم واحد = 2.
9 AKs × 2 ثلاثة توائم = 18.
1 AK 3 ثلاثة توائم = 3.
5 AKs × 4 ثلاثة توائم = 20.
3 AKs × 6 ثلاثة توائم = 18.
إجمالي 61 رمزًا ثلاثيًا لـ 20 حمضًا أمينيًا.
3. وجود علامات الترقيم بين الجينات.
تعريف:
الجين هو جزء من DNA يرمز لسلسلة بولي ببتيد واحدة أو جزيء واحد tPHK, صRNA أوsPHK.
الجيناتtPHK, rPHK, sPHKالبروتينات لا ترميز.
في نهاية كل جين يقوم بتشفير بولي ببتيد ، يوجد واحد على الأقل من ثلاثة توائم ترميز رموز إيقاف RNA ، أو إشارات التوقف. في mRNA تبدو كالتالي: UAA ، UAG ، UGA . يقومون بإنهاء (إنهاء) البث.
تقليديًا ، ينطبق الكودون أيضًا على علامات الترقيمأغسطس - الأول بعد تسلسل الزعيم. (انظر المحاضرة 8) تؤدي وظيفة الحرف الكبير. في هذا الموقف ، يتم ترميز الفورميل ميثيونين (في بدائيات النوى).
4. التفرد.
كل ثلاثة توائم يشفر حمض أميني واحد فقط أو عبارة عن فاصل ترجمة.
الاستثناء هو الكودونأغسطس . في بدائيات النوى ، في الموضع الأول (حرف كبير) يرمز إلى فورميل ميثيونين ، وفي أي موضع آخر يرمز للميثيونين.
5. الضغط ، أو عدم وجود علامات الترقيم داخل الجين.
داخل الجين ، كل نوكليوتيد هو جزء من كودون مهم.
في عام 1961 ، أثبت سيمور بينزر وفرانسيس كريك تجريبياً أن الشفرة ثلاثية ومضغوطة.
جوهر التجربة: الطفرة "+" - إدخال نوكليوتيد واحد. "-" الطفرة - فقدان أحد النوكليوتيدات. طفرة واحدة "+" أو "-" في بداية الجين تفسد الجين بأكمله. كما أن الطفرة المزدوجة "+" أو "-" تفسد الجين بأكمله.
الطفرة الثلاثية "+" أو "-" في بداية الجين تفسد جزءًا فقط منه. الطفرة الرباعية "+" أو "-" تفسد الجين بأكمله مرة أخرى.
التجربة تثبت ذلك الشفرة ثلاثية ولا توجد علامات ترقيم داخل الجين.أجريت التجربة على جينين متجاورين من الملتهمة وأظهرت بالإضافة إلى ذلك ، وجود علامات الترقيم بين الجينات.
6. براعة.
الشفرة الجينية هي نفسها لجميع الكائنات الحية على الأرض.
في عام 1979 افتتح بوريل المثاليرمز الميتوكوندريا البشرية.
تعريف:
"المثالي" هو الكود الجيني الذي يتم فيه استيفاء قاعدة انحطاط الشفرة شبه المزدوجة: إذا تزامن النوكليوتيدات الأولى والثانية في مجموعتين من ثلاثة توائم ، والنيوكليوتيدات الثالثة تنتمي إلى نفس الفئة (كلاهما من البيورينات أو كلاهما بيريميدين) ، ثم هذه الثلاثة توائم ترميز نفس الحمض الأميني.
هناك استثناءان لهذه القاعدة في التعليمات البرمجية العامة. كلا الانحرافين عن الكود المثالي في الكوني مرتبطان بالنقاط الأساسية: بداية ونهاية تخليق البروتين:
كودون | عالمي الرمز | رموز الميتوكوندريا |
|||
الفقاريات | اللافقاريات | خميرة | النباتات |
||
قف | قف |
||||
مع UA | |||||
أ ز أ | قف | ||||
قف | 230 بدائل لا تغير صنف الحمض الأميني المشفر. إلى القابلية للتيسير. في عام 1956 ، اقترح جورجي جاموف نوعًا مختلفًا من الكود المتداخل. وفقًا لرمز Gamow ، فإن كل نوكليوتيد ، بدءًا من الثالث في الجين ، هو جزء من 3 كودونات. عندما تم فك الشفرة الجينية ، اتضح أنها غير متداخلة ، أي كل نوكليوتيد هو جزء من كودون واحد فقط. مزايا الكود الجيني المتداخل: الترابط ، اعتماد أقل لبنية البروتين على إدخال أو حذف نيوكليوتيد. العيب: اعتماد كبير على بنية البروتين على استبدال النوكليوتيدات والقيود على الجيران. في عام 1976 ، تم تسلسل الحمض النووي للعاثية φX174. يحتوي على DNA دائري واحد مجدول مكون من 5375 نيوكليوتيد. كان من المعروف أن العاثية تقوم بتشفير 9 بروتينات. بالنسبة لستة منهم ، تم تحديد الجينات الموجودة واحدة تلو الأخرى. اتضح أن هناك تداخل. الجين E موجود بالكامل داخل الجيند . يظهر كودون البدء الخاص به نتيجة لتحول نيوكليوتيد واحد في القراءة. الجيني يبدأ حيث ينتهي الجيند . كودون بدء الجيناتي يتداخل مع كودون إنهاء الجيند بسبب تحول اثنين من النيوكليوتيدات. يسمى التصميم "تغيير إطار القراءة" بعدد من النيوكليوتيدات التي ليست من مضاعفات الثلاثة. حتى الآن ، تم عرض التداخل فقط لعدد قليل من العاثيات. القدرة المعلوماتية للحمض النووي يوجد 6 مليارات شخص على الأرض. معلومات وراثية عنها 4x10 13 صفحة كتاب. سوف تشغل هذه الصفحات مساحة 6 مباني NSU. 6 × 10 9 الحيوانات المنوية تأخذ نصف كشتبان. يأخذ حمضهم النووي أقل من ربع الكشتبان. | ||||
إذا كان من الضروري تصنيع البروتينات ، تظهر مشكلة خطيرة أمام الخلية - يتم تخزين المعلومات الموجودة في الحمض النووي في شكل تسلسل مشفر 4 أحرف(النيوكليوتيدات) والبروتينات 20 رمزًا مختلفًا(أحماض أمينية). إذا حاولت استخدام جميع الرموز الأربعة دفعة واحدة لتشفير الأحماض الأمينية ، فستحصل فقط على 16 تركيبة ، بينما يوجد 20 من الأحماض الأمينية المكونة للبروتين. لا يكفي ...
وهناك مثال على التفكير العبقري في هذا الصدد:
"خذ ، على سبيل المثال ، مجموعة أوراق اللعب التي نوليها اهتمامًا فقط لنوع البطاقة. كم ثلاثة توائم من نفس النوع يمكن الحصول عليها؟ أربعة بالطبع: ثلاثة من القلوب ، وثلاثة من الماس ، وثلاثة بستوني ، وثلاثة من الهراوات. كم عدد ثلاثة توائم بطاقتين من نفس النوع وواحدة من الأخرى؟ لنفترض أن لدينا أربعة خيارات للبطاقة الثالثة. إذن لدينا 4x3 = 12 احتمالًا. بالإضافة إلى ذلك ، لدينا أربعة توائم بثلاث بطاقات مختلفة. إذن ، 4 + 12 + 4 = 20 ، وهذا هو العدد الدقيق للأحماض الأمينية التي أردنا الحصول عليها "(جورج جامو ، الإنجليزي جورج جامو ، 1904-1968 ، عالم الفيزياء النظرية السوفياتي والأمريكي ، عالم الفيزياء الفلكية وشائع العلوم).
في الواقع ، أظهرت التجارب أنه لكل حمض أميني هناك نوعان من النيوكليوتيدات الإلزامية ومتغير ثالث ، أقل تحديدًا (" تأثير هزازإذا أخذت ثلاثة أحرف من أربعة ، تحصل على 64 مجموعة ، والتي تتداخل بشكل كبير مع عدد الأحماض الأمينية. وهكذا ، وجد أن أي حمض أميني مشفر بثلاثة نيوكليوتيدات. وهذا الثلاثي يسمى كودون. هم ، كما ذكرنا سابقًا ، هناك 64 خيارًا. ثلاثة منهم لا يرمز إلى أي حمض أميني ، هؤلاء هم ما يسمى " أكواد هراء"(فرنسي. غير الحواس- هراء) أو "إيقاف الكودونات".
الكود الجيني
الكود الجيني (البيولوجي) هو طريقة لترميز المعلومات حول بنية البروتينات في شكل تسلسل نيوكليوتيد. وهي مصممة لترجمة لغة النيوكليوتيدات المكونة من أربعة أرقام (A ، G ، U ، C) إلى لغة مكونة من عشرين رقمًا من الأحماض الأمينية. لها سمات مميزة:
- الثلاثيةتشكل ثلاثة نيوكليوتيدات كودون يرمز إلى حمض أميني. هناك 61 كودون حاسة في المجموع.
- النوعية(أو التفرد) - كل كودون يتوافق مع حمض أميني واحد فقط.
- انحطاطيمكن أن يحتوي أحد الأحماض الأمينية على عدة أكواد.
- براعه- الشفرة البيولوجية هي نفسها لجميع أنواع الكائنات الحية على الأرض (ومع ذلك ، هناك استثناءات في الميتوكوندريا للثدييات).
- علاقة خطية متداخلة- يتوافق تسلسل الكودونات مع تسلسل الأحماض الأمينية في البروتين المشفر.
- غير التداخل- ثلاثة توائم لا تتداخل مع بعضها البعض ، وتقع جنبًا إلى جنب.
- علامات الترقيم مفقودة- لا توجد نيوكليوتيدات إضافية أو أي إشارات أخرى بين التوائم الثلاثة.
- أحادي الاتجاه- أثناء تخليق البروتين ، تستمر قراءة الكودون بالتتابع ، بدون ثغرات أو تراجع.
ومع ذلك ، فمن الواضح أن الكود البيولوجي لا يمكن أن يعبر عن نفسه بدون جزيئات إضافية تؤدي وظيفة انتقالية أو وظيفة المحول.
دور محول نقل الحمض النووي الريبي
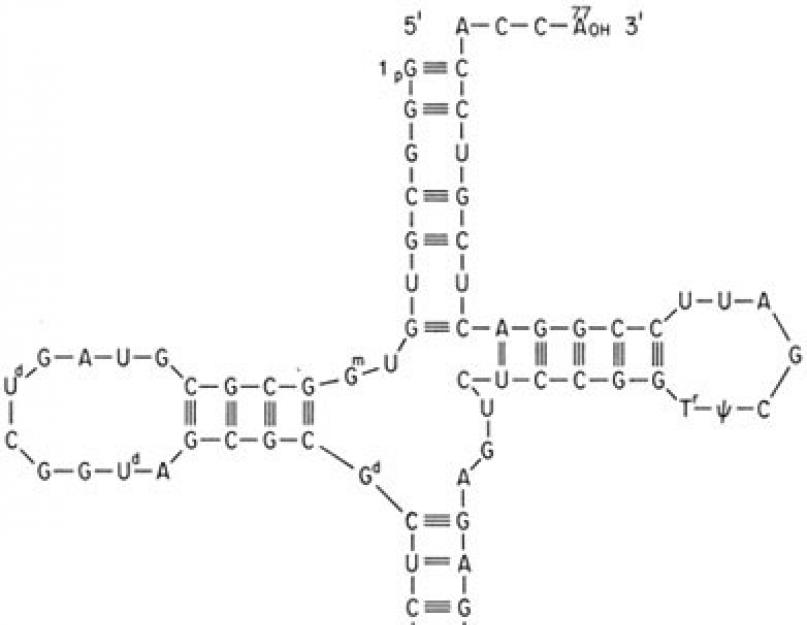
نقل الحمض النووي الريبي هو الوسيط الوحيد بين تسلسل الحمض النووي المكون من 4 أحرف وتسلسل البروتين المكون من 20 حرفًا.
يحتوي كل نقل RNA على تسلسل ثلاثي محدد في حلقة anticodon ( أنتيكودون) ويمكنه فقط إرفاق حمض أميني يطابق هذا المضاد. إن وجود واحد أو آخر من مضادات الكودون في الحمض النووي الريبي هو الذي يحدد الحمض الأميني الذي سيتم تضمينه في جزيء البروتين ، tk. لا الريبوسوم ولا الرنا المرسال يتعرفان على الأحماض الأمينية.
في هذا الطريق، دور محول من الحمض الريبي النووي النقالهو:
- في ارتباط محدد للأحماض الأمينية ،
- على وجه التحديد ، وفقًا لتفاعل الكودون-أنتي كودون ، المرتبط بـ mRNA ،
- ونتيجة لذلك ، في إدراج الأحماض الأمينية في سلسلة البروتين وفقًا لمعلومات الرنا المرسال.
يتم تنفيذ ارتباط الحمض الأميني بـ tRNA بواسطة إنزيم aminoacyl-tRNA synthetase، والتي لها خصوصية لمركبين في وقت واحد: أي حمض أميني و tRNA المقابل له. يتطلب التفاعل رابطتين كبيرتين من ATP. يرتبط الحمض الأميني بنهاية 3 'من حلقة مستقبل الحمض الريبي النووي النقال من خلال مجموعة α-carboxyl ، وتصبح الرابطة بين الحمض الأميني و tRNA ماكرو. تظل مجموعة α-amino مجانية.
تفاعل تخليق Aminoacyl-tRNA
نظرًا لوجود حوالي 60 tRNAs مختلفًا ، فإن بعض الأحماض الأمينية تتوافق مع اثنين أو أكثر من tRNAs. تسمى الحمض الريبي النووي النقال المختلف الذي يربط نفس الحمض الأميني مستقبلات متساوية.
في عملية التمثيل الغذائي في الجسم دور قيادي
ينتمي إلى البروتينات والأحماض النووية.
تشكل المواد البروتينية أساس جميع الهياكل الحيوية للخلايا ، ولها تفاعل عالي بشكل غير عادي ، وتتمتع بوظائف تحفيزية.
تعد الأحماض النووية جزءًا من أهم عضو في الخلية - النواة ، بالإضافة إلى السيتوبلازم والريبوزومات والميتوكوندريا وما إلى ذلك. تلعب الأحماض النووية دورًا أساسيًا مهمًا في الوراثة وتنوع الجسم وتخليق البروتين.
يخططتركيب يتم تخزين البروتين في نواة الخلية ، ويحدث التركيب المباشر خارج النواة ، لذلك فهو ضروري خدمة التوصيلمشفر خطة من النواة إلى موقع التوليف. يتم تنفيذ خدمة التوصيل هذه بواسطة جزيئات RNA.
تبدأ العملية في جوهر الخلايا: جزء من "سلم" الحمض النووي ينفتح ويفتح. نتيجة لذلك ، تشكل أحرف RNA روابط مع أحرف DNA المفتوحة لإحدى خيوط DNA. يقوم الإنزيم بنقل أحرف RNA لربطها في خيط. لذلك تتم إعادة كتابة أحرف الحمض النووي في أحرف RNA. يتم فصل سلسلة RNA المشكلة حديثًا ، ويلتوى "سلم" الحمض النووي مرة أخرى. تسمى عملية قراءة المعلومات من الحمض النووي وتوليف قالب الحمض النووي الريبي الخاص به النسخ ، ويسمى الحمض النووي الريبي المركب بالمعلومات أو ط- RNA .
بعد مزيد من التعديلات ، يكون هذا النوع من mRNA المشفر جاهزًا. ط- RNA يخرج من النواةويذهب إلى موقع تخليق البروتين ، حيث يتم فك رموز الأحرف i-RNA. كل مجموعة من ثلاثة أحرف من i-RNA تشكل "حرفًا" يرمز إلى حمض أميني معين.
نوع آخر من الحمض النووي الريبي يبحث عن هذا الحمض الأميني ، يلتقطه بمساعدة إنزيم ، ويوصله إلى موقع تخليق البروتين. يسمى هذا RNA نقل RNA ، أو tRNA. أثناء قراءة رسالة mRNA وترجمتها ، تنمو سلسلة الأحماض الأمينية. تلتف هذه السلسلة وتطوي في شكل فريد ، مكونة نوعًا واحدًا من البروتين. حتى عملية طي البروتين رائعة: استخدام الكمبيوتر لحساب كل شيء والخياراتسوف يستغرق الأمر 1027 سنة (!) لطي بروتين متوسط الحجم يتكون من 100 حمض أميني. ولتشكيل سلسلة من 20 حمضًا أمينيًا في الجسم ، لا تستغرق أكثر من ثانية واحدة ، وتحدث هذه العملية بشكل مستمر في جميع خلايا الجسم.
الجينات والشفرة الجينية وخصائصها.
يعيش حوالي 7 مليارات شخص على الأرض. باستثناء 25-30 مليون زوج من التوائم المتماثلة ، ثم جينيا كل الناس مختلفون : كل منها فريد من نوعه ، له خصائص وراثية فريدة ، سمات شخصية ، قدرات ، مزاج.
يتم شرح هذه الاختلافات الاختلافات في الأنماط الجينية- مجموعات من جينات الكائن الحي ؛ كل واحد فريد. تتجسد السمات الجينية لكائن معين في البروتينات - وبناءً على ذلك ، تختلف بنية البروتين لدى شخص ما ، على الرغم من اختلافها قليلاً ، عن بروتين شخص آخر.
هذا ليس ما اعنيهأن البشر لا يمتلكون نفس البروتينات بالضبط. قد تكون البروتينات التي تؤدي نفس الوظائف هي نفسها أو تختلف اختلافًا طفيفًا جدًا بواحد أو اثنين من الأحماض الأمينية عن بعضها البعض. ولكن غير موجود على الأرض من البشر (باستثناء التوائم المتطابقة) ، حيث ستكون جميع البروتينات هي نفسها .
معلومات حول التركيب الأساسي للبروتينمشفرة كسلسلة من النيوكليوتيدات في جزء من جزيء الحمض النووي ، الجين - وحدة المعلومات الوراثية للكائن الحي. يحتوي كل جزيء DNA على العديد من الجينات. يتكون مجموع كل جينات الكائن الحي من الطراز العرقى . في هذا الطريق،
الجين هو وحدة المعلومات الوراثية للكائن الحي ، والتي تتوافق مع قسم منفصل من الحمض النووي
يتم ترميز المعلومات الوراثية باستخدام الكود الجيني ، وهو عالمي لجميع الكائنات الحية ويختلف فقط في تناوب النيوكليوتيدات التي تشكل الجينات وترمز لبروتينات كائنات معينة.
الكود الجيني يتكون من ثلاثة توائم (ثلاثة توائم) من نيوكليوتيدات الحمض النووي ، مجتمعة في تسلسلات مختلفة (AAT ، HCA ، ACG ، THC ، إلخ) ، كل منها يشفر حمض أميني معين (والذي سيتم بناؤه في سلسلة البولي ببتيد).
في الحقيقة الشفرة
العد تسلسل النيوكليوتيدات في جزيء i-RNA
، لان يزيل المعلومات من الحمض النووي (العملية النسخ
) ويترجمها إلى سلسلة من الأحماض الأمينية في جزيئات البروتينات المركبة (عملية البث
).
يتضمن تكوين mRNA النيوكليوتيدات A-C-G-U ، والتي تسمى ثلاثة توائم منها الكودونات
: سيصبح الثلاثي DNA CHT على mRNA ثلاثي HCA ، وسيصبح ثلاثي AAG DNA ثلاثي UUC. بالضبط أكواد i-RNA
يعكس الشفرة الجينية في السجل.
في هذا الطريق، الكود الجيني - نظام موحد لتسجيل المعلومات الوراثية في جزيئات الحمض النووي في شكل سلسلة من النيوكليوتيدات . يعتمد الكود الجيني على استخدام أبجدية تتكون من أربعة أحرف نوكليوتيد فقط تختلف في القواعد النيتروجينية: A ، T ، G ، C.
الخصائص الرئيسية للشفرة الجينية:

1. الكود الجيني ثلاثة توائم. الثلاثي (الكودون) هو سلسلة من ثلاثة نيوكليوتيدات ترمز لحمض أميني واحد. نظرًا لأن البروتينات تحتوي على 20 نوعًا من الأحماض الأمينية ، فمن الواضح أنه لا يمكن ترميز كل منها بواسطة نيوكليوتيد واحد ( نظرًا لوجود أربعة أنواع فقط من النيوكليوتيدات في الحمض النووي ، في هذه الحالة يبقى 16 حمضًا أمينيًا غير مشفر). كما أن اثنين من النيوكليوتيدات لترميز الأحماض الأمينية لا يكفيان أيضًا ، لأنه في هذه الحالة يمكن تشفير 16 فقط من الأحماض الأمينية. هذا يعني أن أقل عدد من النيوكليوتيدات يشفر حمض أميني واحد يجب أن يكون ثلاثة على الأقل. في هذه الحالة ، يكون عدد توائم النوكليوتيدات الثلاثية الممكنة هو 43 = 64.
2. التكرار (انحلال)الكود هو نتيجة لطبيعته الثلاثية ويعني أن حمض أميني واحد يمكن ترميزه بعدة ثلاثة توائم (حيث يوجد 20 حمضًا أمينيًا ، وهناك 64 ثلاثيًا) ، باستثناء الميثيونين والتربتوفان ، والتي يتم ترميزها بواسطة واحد فقط ثلاثة توائم. بالإضافة إلى ذلك ، تؤدي بعض التوائم الثلاثة وظائف محددة: في جزيء mRNA ، تقوم الثلاثية UAA و UAG و UGA بإنهاء الكودونات ، أي قف- الإشارات التي توقف تخليق سلسلة البولي ببتيد. الثلاثي المقابل للميثيونين (AUG) ، الذي يقف في بداية سلسلة الحمض النووي ، لا يشفر الحمض الأميني ، ولكنه يؤدي وظيفة بدء القراءة (المثيرة).
3. غموض الكود - إلى جانب التكرار ، يمتلك الرمز الخاصية التفرد : كل كودون يطابق فقط واحدحمض أميني محدد.
4. علاقة خطية متداخلة الكود ، أي تسلسل النيوكليوتيدات في الجين بالضبطيتوافق مع تسلسل الأحماض الأمينية في البروتين.
5. الكود الجيني غير متداخلة ومضغوطة ، أي لا يحتوي على "علامات الترقيم". هذا يعني أن عملية القراءة لا تسمح بإمكانية تداخل الأعمدة (ثلاثة توائم) ، وبدءًا من كودون معين ، تستمر القراءة ثلاث مرات بثلاثة أضعاف حتى قف-إشارات ( رموز الإنهاء).
6. الكود الجيني عالمي ، أي أن الجينات النووية لجميع الكائنات الحية تقوم بتشفير المعلومات حول البروتينات بنفس الطريقة ، بغض النظر عن مستوى التنظيم والموقع المنهجي لهذه الكائنات.
يوجد جداول الكود الجيني لفك التشفير الكودونات i-RNA وسلاسل بناء من جزيئات البروتين.

تفاعلات تركيب المصفوفة.
في الأنظمة الحية ، توجد ردود أفعال غير معروفة في الطبيعة غير الحية - تفاعلات تركيب المصفوفة.
مصطلح "المصفوفة"في التكنولوجيا ، يشيرون إلى الشكل المستخدم في صب العملات المعدنية والميداليات ونوع الطباعة: يستنسخ المعدن المقوى بالضبط جميع تفاصيل النموذج المستخدم في الصب. توليف المصفوفةيشبه الصب على مصفوفة: يتم تصنيع الجزيئات الجديدة بما يتفق بدقة مع الخطة الموضوعة في بنية الجزيئات الموجودة بالفعل.
يكمن مبدأ المصفوفة في الصميمأهم التفاعلات التركيبية للخلية ، مثل تخليق الأحماض النووية والبروتينات. في هذه التفاعلات ، يتم توفير تسلسل دقيق ومحدد بدقة للوحدات الأحادية في البوليمرات المركبة.
هذا هو حيث الاتجاه سحب المونومرات إلى موقع محددالخلايا - إلى جزيئات تعمل كمصفوفة حيث يحدث التفاعل. إذا حدثت مثل هذه التفاعلات نتيجة تصادم عشوائي للجزيئات ، فستتقدم ببطء إلى ما لا نهاية. يتم تركيب الجزيئات المعقدة على أساس مبدأ المصفوفة بسرعة وبدقة. دور المصفوفة تلعب الجزيئات الكبيرة للأحماض النووية في تفاعلات المصفوفة DNA أو RNA .

جزيئات أحادية، التي يتم تصنيع البوليمر منها - النيوكليوتيدات أو الأحماض الأمينية - وفقًا لمبدأ التكامل يتم ترتيبها وتثبيتها على المصفوفة بترتيب محدد بدقة ومحددة مسبقًا.
ثم يأتي "التشابك" لوحدات المونومر في سلسلة بوليمر، ويتم إسقاط البوليمر النهائي من المصفوفة.
بعد ذلك المصفوفة جاهزةلتجميع جزيء بوليمر جديد. من الواضح أنه مثل عملة واحدة فقط ، يمكن صب حرف واحد على قالب معين ، لذلك يمكن "تجميع" بوليمر واحد فقط على جزيء مصفوفة معين.
نوع مصفوفة من ردود الفعل- سمة محددة لكيمياء النظم الحية. إنها أساس الملكية الأساسية لجميع الكائنات الحية - قدرتها على إعادة إنتاج نوعها.
تفاعلات تركيب المصفوفة
1. تكرار الحمض النووي - النسخ المتماثل (من خط الطول. النسخ المتماثل - التجديد) - عملية تخليق جزيء ابنة من حمض الديوكسي ريبونوكلييك على مصفوفة جزيء الحمض النووي الأصل. أثناء الانقسام اللاحق للخلية الأم ، تتلقى كل خلية ابنة نسخة واحدة من جزيء DNA مطابق للحمض النووي للخلية الأم الأصلية. تضمن هذه العملية النقل الدقيق للمعلومات الجينية من جيل إلى جيل. يتم إجراء تكرار الحمض النووي بواسطة مركب إنزيم معقد ، يتكون من 15-20 بروتينًا مختلفًا ، يُسمى ريبيزوم . مادة التوليف هي نيوكليوتيدات حرة موجودة في سيتوبلازم الخلايا. يكمن المعنى البيولوجي للتكاثر في النقل الدقيق للمعلومات الوراثية من الجزيء الأصل إلى الجزيء الابنة ، والذي يحدث عادةً أثناء انقسام الخلايا الجسدية.
يتكون جزيء الحمض النووي من خيطين متكاملين. ترتبط هذه السلاسل ببعضها البعض بواسطة روابط هيدروجينية ضعيفة يمكن أن تنكسر بواسطة الإنزيمات. جزيء الحمض النووي قادر على المضاعفة الذاتية (النسخ المتماثل) ، ويتم تصنيع نصف جديد منه على كل نصف قديم من الجزيء.
بالإضافة إلى ذلك ، يمكن تصنيع جزيء mRNA على جزيء DNA ، والذي يقوم بعد ذلك بنقل المعلومات الواردة من DNA إلى موقع تخليق البروتين.
يتبع نقل المعلومات وتخليق البروتين مبدأ المصفوفة ، يمكن مقارنته بعمل المطبعة في المطبعة. يتم نسخ المعلومات من الحمض النووي مرارًا وتكرارًا. إذا حدثت أخطاء أثناء النسخ ، فسوف تتكرر في جميع النسخ اللاحقة.

صحيح أن بعض الأخطاء في نسخ المعلومات بواسطة جزيء DNA يمكن تصحيحها - تسمى عملية إزالة الأخطاء تعويضات. أول التفاعلات في عملية نقل المعلومات هو تكرار جزيء الحمض النووي وتخليق خيوط DNA الجديدة.
2. النسخ (من النسخ اللاتينية - إعادة الكتابة) - عملية تخليق الحمض النووي الريبي باستخدام الحمض النووي كقالب ، والتي تحدث في جميع الخلايا الحية. بمعنى آخر ، إنه نقل المعلومات الجينية من الحمض النووي إلى الحمض النووي الريبي.
يتم تحفيز النسخ بواسطة إنزيم بوليميراز الحمض النووي الريبي المعتمد على الحمض النووي. يتحرك بوليميراز الحمض النووي الريبي على طول جزيء الحمض النووي في الاتجاه 3 "→ 5". يتكون النسخ من خطوات البدء والاستطالة والإنهاء . وحدة النسخ هي الأوبرا ، وهو جزء من جزيء الحمض النووي يتكون من المروج ، والجزء المكتوب ، والمنهي . يتكون i-RNA من خيط واحد ويتم تصنيعه على DNA وفقًا لقاعدة التكامل بمشاركة إنزيم ينشط بداية ونهاية تخليق جزيء i-RNA.
يدخل جزيء الرنا المرسال النهائي إلى السيتوبلازم على الريبوسومات ، حيث يتم تركيب سلاسل البولي ببتيد.
3. إذاعة (من اللات. ترجمة- النقل ، الحركة) - عملية تخليق البروتين من الأحماض الأمينية على مصفوفة المعلوماتية (المصفوفة) RNA (mRNA ، mRNA) التي يقوم بها الريبوسوم. بمعنى آخر ، هذه هي عملية ترجمة المعلومات الواردة في تسلسل النوكليوتيدات لـ i-RNA إلى تسلسل الأحماض الأمينية في عديد الببتيد.

4. النسخ العكسي هي عملية تكوين DNA مزدوج الشريطة بناءً على معلومات من الحمض النووي الريبي أحادي الشريطة. تسمى هذه العملية النسخ العكسي ، حيث يحدث نقل المعلومات الجينية في الاتجاه "العكسي" بالنسبة إلى النسخ. كانت فكرة النسخ العكسي في البداية غير مرغوبة للغاية ، لأنها تتعارض مع العقيدة المركزية للبيولوجيا الجزيئية ، التي افترضت أن الحمض النووي يُنسخ إلى RNA ثم يُترجم إلى بروتينات.
 ومع ذلك ، في عام 1970 ، اكتشف كل من Temin و Baltimore بشكل مستقل إنزيمًا يسمى نسخة عكسية (رجوع)
، وتم التأكيد أخيرًا على إمكانية النسخ العكسي. في عام 1975 ، حصل كل من Temin و Baltimore على جائزة نوبل في علم وظائف الأعضاء أو الطب. بعض الفيروسات (مثل فيروس نقص المناعة البشرية الذي يسبب عدوى فيروس نقص المناعة البشرية) لديها القدرة على نسخ الحمض النووي الريبي إلى الحمض النووي. فيروس نقص المناعة البشرية لديه جينوم الحمض النووي الريبي الذي يندمج في الحمض النووي. نتيجة لذلك ، يمكن دمج الحمض النووي للفيروس مع جينوم الخلية المضيفة. يسمى الإنزيم الرئيسي المسؤول عن تخليق الحمض النووي من الحمض النووي الريبي العودة. إحدى وظائف الانعكاس هي الخلق الحمض النووي التكميلي
(كدنا) من الجينوم الفيروسي. إنزيم الريبونوكلياز المرتبط به يشق الحمض النووي الريبي ، ويصنع إنزيم الريكسيتاز (كدنا) من الحلزون المزدوج للحمض النووي. تم دمج (كدنا) في جينوم الخلية المضيفة عن طريق التكامل. النتيجه هي تخليق البروتينات الفيروسية بواسطة الخلية المضيفةالتي تشكل فيروسات جديدة. في حالة فيروس نقص المناعة البشرية ، يتم أيضًا برمجة موت الخلايا المبرمج (موت الخلايا) للخلايا اللمفاوية التائية. في حالات أخرى ، قد تظل الخلية موزعًا للفيروسات.
ومع ذلك ، في عام 1970 ، اكتشف كل من Temin و Baltimore بشكل مستقل إنزيمًا يسمى نسخة عكسية (رجوع)
، وتم التأكيد أخيرًا على إمكانية النسخ العكسي. في عام 1975 ، حصل كل من Temin و Baltimore على جائزة نوبل في علم وظائف الأعضاء أو الطب. بعض الفيروسات (مثل فيروس نقص المناعة البشرية الذي يسبب عدوى فيروس نقص المناعة البشرية) لديها القدرة على نسخ الحمض النووي الريبي إلى الحمض النووي. فيروس نقص المناعة البشرية لديه جينوم الحمض النووي الريبي الذي يندمج في الحمض النووي. نتيجة لذلك ، يمكن دمج الحمض النووي للفيروس مع جينوم الخلية المضيفة. يسمى الإنزيم الرئيسي المسؤول عن تخليق الحمض النووي من الحمض النووي الريبي العودة. إحدى وظائف الانعكاس هي الخلق الحمض النووي التكميلي
(كدنا) من الجينوم الفيروسي. إنزيم الريبونوكلياز المرتبط به يشق الحمض النووي الريبي ، ويصنع إنزيم الريكسيتاز (كدنا) من الحلزون المزدوج للحمض النووي. تم دمج (كدنا) في جينوم الخلية المضيفة عن طريق التكامل. النتيجه هي تخليق البروتينات الفيروسية بواسطة الخلية المضيفةالتي تشكل فيروسات جديدة. في حالة فيروس نقص المناعة البشرية ، يتم أيضًا برمجة موت الخلايا المبرمج (موت الخلايا) للخلايا اللمفاوية التائية. في حالات أخرى ، قد تظل الخلية موزعًا للفيروسات.
يمكن تمثيل تسلسل تفاعلات المصفوفة أثناء التخليق الحيوي للبروتين كرسم تخطيطي.

في هذا الطريق، التخليق الحيوي للبروتين- هذا أحد أنواع التبادل البلاستيكي ، حيث تتحقق المعلومات الوراثية المشفرة في جينات الحمض النووي في تسلسل معين من الأحماض الأمينية في جزيئات البروتين.
جزيئات البروتين هي في الأساس سلاسل بولي ببتيدتتكون من الأحماض الأمينية الفردية. لكن الأحماض الأمينية ليست نشطة بما يكفي للتواصل مع بعضها البعض من تلقاء نفسها. لذلك ، قبل الاتصال ببعضها البعض وتشكيل جزيء البروتين ، يجب أن تكون الأحماض الأمينية تفعيل . يحدث هذا التنشيط تحت تأثير إنزيمات خاصة.
نتيجة للتنشيط ، يصبح الحمض الأميني أكثر قابلية للتغير ، وتحت تأثير نفس الإنزيم ، يرتبط بـ t- RNA. يتوافق كل حمض أميني مع عنصر t محدد بدقة RNA، والتي تجد "لها" الأحماض الأمينية و يتحملفي الريبوسوم.
لذلك ، يتلقى الريبوسوم مختلفًا تنشيط الأحماض الأمينية المرتبطة بهمر- RNA. الريبوسوم مثل ناقللتجميع سلسلة بروتين من مختلف الأحماض الأمينية التي تدخلها.
بالتزامن مع t-RNA ، حيث "يجلس" الحمض الأميني الخاص به ، " الإشارة»من الحمض النووي الموجود في النواة. وفقًا لهذه الإشارة ، يتم تصنيع بروتين أو آخر في الريبوسوم.
لا يتم تنفيذ التأثير التوجيهي للحمض النووي على تخليق البروتين بشكل مباشر ، ولكن بمساعدة وسيط خاص - مصفوفةأو رسول RNA (مرناأو ط- RNA)، أيّ توليفها في النواةلا يتأثر بالحمض النووي ، لذا فإن تركيبته تعكس تكوين الحمض النووي. جزيء الحمض النووي الريبي ، كما كان ، هو قالب من شكل الحمض النووي. يدخل mRNA المركب الريبوسوم وينقله ، كما كان ، إلى هذا الهيكل خطة- في أي ترتيب يجب أن تتحد الأحماض الأمينية المنشطة التي تدخل الريبوسوم مع بعضها البعض من أجل تخليق بروتين معين. خلاف ذلك، يتم نقل المعلومات الجينية المشفرة في الحمض النووي إلى mRNA ثم إلى البروتين.
يدخل جزيء mRNA الريبوسوم و ومضاتلها. يتم تحديد ذلك الجزء الموجود حاليًا في الريبوسوم كودون (ثلاثي)، يتفاعل بطريقة محددة تمامًا مع بنية مناسبة لها ثلاثي (أنتيكودون)في نقل الحمض النووي الريبي الذي جلب الحمض الأميني إلى الريبوسوم.
يقترب نقل الحمض النووي الريبي بحمضه الأميني من كودون معين من الرنا المرسال و يربطمعه إلى الموقع التالي المجاور لـ i-RNA ينضم إلى الحمض الريبي النووي النقال آخر مع حمض أميني مختلفوهكذا حتى يتم قراءة سلسلة i-RNA بأكملها ، حتى يتم ربط جميع الأحماض الأمينية بالترتيب المناسب ، لتشكيل جزيء بروتين. و t-RNA ، الذي أوصل الحمض الأميني إلى موقع معين من سلسلة البولي ببتيد ، تحرر من الأحماض الأمينيةويخرج الريبوسوم.
ثم مرة أخرى في السيتوبلازم ، يمكن للحمض الأميني المرغوب أن ينضم إليه ، وسوف ينقله مرة أخرى إلى الريبوسوم. في عملية تخليق البروتين ، لا تشارك ريبوسومات واحدة ، بل عدة ريبوسومات ، متعددة الريبوسومات ، في نفس الوقت.
المراحل الرئيسية لنقل المعلومات الجينية:
1. توليف على الحمض النووي كما في قالب mRNA (النسخ)
2. توليف سلسلة البولي ببتيد في الريبوسومات وفقًا للبرنامج الوارد في i-RNA (ترجمة)
.
المراحل عالمية لجميع الكائنات الحية ، لكن العلاقات الزمانية والمكانية لهذه العمليات تختلف في البدائل وحقيقيات النوى.
في بدائيات النوىيمكن أن يحدث النسخ والترجمة في وقت واحد لأن الحمض النووي يقع في السيتوبلازم. في حقيقيات النوىيتم فصل النسخ والترجمة بشكل صارم في المكان والزمان: يحدث تخليق مختلف RNAs في النواة ، وبعد ذلك يجب أن تغادر جزيئات الحمض النووي الريبي النواة ، مروراً بالغشاء النووي. ثم يتم نقل الحمض النووي الريبي في السيتوبلازم إلى موقع تخليق البروتين.



