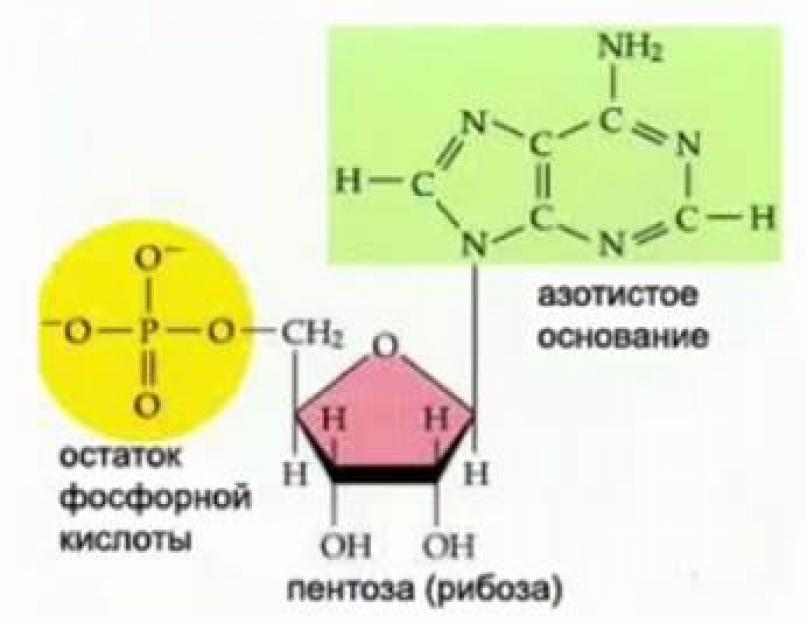
Nukleotidų sudėtyje esantys RNR monomerai turi penkių anglies cukrų (pentozę), fosforo rūgštį (fosforo rūgšties liekaną) ir azoto bazę (žr. 2 pav.).

Ryžiai. 2. RNR nukleotido sandara
Azotinės RNR bazės yra uracilas, citozinas, adeninas ir guaninas. RNR nukleotido monosacharidas pavaizduotas riboze (žr. 2 pav.).
RNR yra viengrandė molekulė, daug mažesnė už DNR molekulę.
RNR molekulėje yra nuo 75 iki 10 000 nukleotidų.

Ryžiai. 3. RNR virusas
Daugelyje virusų, tokių kaip gripo virusas, vienintelė nukleino rūgštis yra RNR molekulė (žr. 3 pav.). Žmonėms patogeniškų virusų, turinčių RNR, yra daugiau nei turinčių DNR. Jie sukelia poliomielitą, hepatitą A, ūmias peršalimo ligas.
Arbovirusai yra nariuotakojų pernešami virusai. Jie yra erkinio ir japoninio encefalito, taip pat geltonosios karštinės sukėlėjai.
Reovirusai (žr. 4 pav.), reti žmogaus kvėpavimo takų ir žarnyno ligų sukėlėjai, sulaukė ypatingo mokslinio susidomėjimo dėl to, kad jų genetinė medžiaga pateikiama dvigrandės RNR molekulės pavidalu.

Ryžiai. 4. Reoviruso struktūra
Taip pat yra retrovirusų, sukeliančių daugybę vėžio formų.
Priklausomai nuo struktūros ir atliekamos funkcijos, išskiriami trys pagrindiniai RNR tipai: ribosominė, transportinė ir informacinė (matrica).
1. Messenger RNR
Tyrimai parodė, kad pasiuntinio RNR sudaro 3-5% viso RNR kiekio ląstelėje. Tai vienagrandė molekulė, kuri susidaro transkripcijos metu vienoje iš DNR molekulės grandinių. Taip yra dėl to, kad DNR branduoliniuose organizmuose yra išsidėsčiusi branduolyje, o baltymų sintezė vyksta ant ribosomų citoplazmoje, todėl atsirado „tarpininko“ poreikis. Messenger RNR atlieka „tarpininko“ funkciją, perduoda informaciją apie baltymo struktūrą iš ląstelės branduolio, kuriame yra DNR, į ribosomas, kur ši informacija realizuojama (žr. 5 pav.).

Ryžiai. 5. Messenger RNR (mRNR)
Priklausomai nuo kopijuojamos informacijos kiekio, pasiuntinio RNR molekulė gali būti skirtingo ilgio.
Dauguma pasiuntinių RNR ląstelėje egzistuoja trumpą laiką. Bakterijų ląstelėse tokių RNR buvimas nustatomas minutėmis, o žinduolių ląstelėse (eritrocituose) hemoglobino (baltymų) sintezė tęsiasi kelias dienas, kai eritrocitai netenka branduolio.
2. Ribosominė RNR
Ribosominės RNR (žr. 6 pav.) sudaro 80 % visų ribosomų, esančių ląstelėje. Šios RNR sintetinamos branduolyje, o ląstelėje yra citoplazmoje, kur kartu su baltymais sudaro ribosomas. Baltymų sintezė vyksta ribosomose. Čia „kodas“, esantis pasiuntinio RNR, paverčiamas baltymo molekulės aminorūgščių seka.

Ryžiai. 6. Ribosominė RNR (rRNR)
3. Perkelkite RNR
Pernešimo RNR (žr. 7 pav.) susidaro DNR esančiame branduolyje ir tada patenka į citoplazmą.

Ryžiai. 7. Perkelkite RNR (tRNR)
Šios RNR sudaro apie 10% viso RNR kiekio ląstelėje. Jie turi trumpiausias 80-100 nukleotidų molekules.
Pernešančios RNR prie savęs pritvirtina aminorūgštį ir perneša ją į baltymų sintezės vietą, į ribosomas.
Visos žinomos pernešančios RNR dėl komplementarios azotinių bazių sąveikos sudaro antrinę struktūrą, savo forma primenančią dobilo lapą (žr. 8 pav.). tRNR molekulė turi dvi aktyvias vietas – viename gale yra tripletas antikodonas, o kitame – akceptoriaus vieta, kuri prijungia aminorūgštį.

Ryžiai. 8. tRNR struktūra („dobilo lapas“)
Kiekviena aminorūgštis atitinka trijų nukleotidų derinį, kuris vadinamas trynukas.

Ryžiai. 9. Genetinio kodo lentelė
Tripletai, koduojantys aminorūgštis kodonai DNR (žr. 9 pav.) – perduodama iRNR tripletų (kodonų) informacijos forma. tRNR dobilo lapo viršuje yra nukleotidų tripletas, kuris yra komplementarus atitinkamam mRNR kodonui (žr. 10 pav.). Šis tripletas skiriasi tRNR, kurios turi skirtingas aminorūgštis, ir koduoja būtent tą aminorūgštį, kurią neša ši tRNR. Jis gavo vardą antikodonas.

Ryžiai. 10. tRNR
Akceptoriaus galas yra tam tikros aminorūgšties „nusileidimo aikštelė“.
Taigi, įvairūs RNR tipai yra viena funkcinė sistema, skirta paveldimos informacijos įgyvendinimui baltymų sintezės būdu.
RNR pasaulio samprata yra ta, kad kadaise RNR molekulė galėjo atlikti ir DNR molekulės, ir baltymų funkciją.
Gyvuose organizmuose beveik visi procesai vyksta dėl baltymų fermentų. Tačiau baltymai negali savaime daugintis ir yra sintetinami į ląsteles remiantis DNR saugoma informacija. Tačiau DNR dubliavimasis vyksta tik dėl baltymų ir RNR dalyvavimo. Vadinasi, susidaro užburtas ratas, dėl kurio gyvybės kilmės teorijos rėmuose spontaniškas tokios sudėtingos sistemos atsiradimas mažai tikėtinas.
Devintojo dešimtmečio pradžioje mokslininkų Cheko ir Altmano (Nobelio chemijos premijos laureatų) laboratorijoje JAV buvo atrastas RNR katalizinis gebėjimas. RNR katalizatoriai buvo pavadinti ribozimai(Žr. 11 pav.).

Ryžiai. 11. RNR ribozimomolekulės, atliekančios katalizės funkciją, struktūra
Paaiškėjo, kad aktyviame ribosomų centre yra ir didelis kiekis ribosominės RNR. RNR taip pat gali sudaryti dvigubą grandinę ir daugintis. Tai reiškia, kad RNR gali egzistuoti visiškai autonomiškai, katalizuojanti medžiagų apykaitos reakcijas, tokias kaip naujų ribonukleatidų sintezė, ir savaime dauginantis, išlaikant katalizines savybes iš kartos į kartą. Dėl atsitiktinių mutacijų kaupimosi atsirado RNR, kurios katalizuoja tam tikrų baltymų sintezę, kurios yra efektyvesni katalizatoriai, todėl šios mutacijos buvo fiksuotos natūralios atrankos metu. Atsirado ir specializuotos genetinės informacijos saugyklos – DNR molekulė, o RNR tapo tarpininku tarp DNR ir baltymų.
Bibliografija
- Kamensky A.A., Kriksunov E.A., Pasechnik V.V. Bendroji biologija 10-11 klasė Bustard, 2005 m.
- Biologija. 10 klasė. Bendroji biologija. Pagrindinis lygis / P.V. Iževskis, O.A. Kornilova, T.E. Loshchilin ir kiti - 2 leidimas, pataisytas. - Ventana-Graf, 2010. - 224 psl.
- Belyajevas D.K. Biologijos 10-11 kl. Bendroji biologija. Pagrindinis lygis. – 11 leid., stereotipas. - M.: Švietimas, 2012. - 304 p.
- Agafonova I.B., Zakharova E.T., Sivoglazovas V.I. Biologijos 10-11 kl. Bendroji biologija. Pagrindinis lygis. - 6 leidimas, pridėti. - Bustard, 2010. - 384 p.
- Orgchem.ru ().
- Appteka.ru ().
- Youtube.com ().
Namų darbai
- 4, 5 klausimai 12 pastraipos pabaigoje (p. 52) – Kamensky A.A., Kriksunov E.A., Pasechnik V.V. „Bendroji biologija“, 10–11 klasė ()
- Kur ląstelėje yra nukleorūgštys?
Ribonukleino rūgštis yra purino ir pirimidino ribonukleotidų kopolimeras, sujungtas vienas su kitu, kaip ir DNR, fosfodiesterio tilteliais (37.6 pav.). Nors šios dvi nukleino rūgščių rūšys turi daug bendro, jos skiriasi viena nuo kitos įvairiais būdais.
1. RNR angliavandenių liekana, prie kurios yra prijungtos purino arba pirimidino bazės ir fosfato grupės, yra ribozė, o ne 2-dezoksiribozė (kaip DNR).
2. RNR pirimidino komponentai skiriasi nuo DNR komponentų. RNR, taip pat DNR sudėtis apima adenino, guanino ir citozino nukleotidus. Tuo pačiu metu RNR (išskyrus kai kuriuos specialius atvejus, kuriuos aptarsime toliau) nėra timino, jo vietą RNR molekulėje užima uracilas.
3. RNR yra viengrandė molekulė (skirtingai nei DNR, kurios struktūra yra dvigrandė), tačiau jei RNR grandinėje yra sekcijų su komplementaria seka (priešingu poliškumu), viena RNR grandinė gali susilankstyti ir susiformuoti -vadinamos „plaukų segtukais“, konstrukcijos, turinčios dvisruoges charakteristikas (37.7 pav.).

Ryžiai. 37.6. Ribonukleorūgšties (RNR) molekulės fragmentas, kuriame purino ir pirimidino bazės - adeninas (A), uracilas (U), citozinas (C) ir guaninas (- yra sulaikomos fosfodiesterio pagrindo, jungiančio ribozilo liekanas, sujungtas N- glikozidinis ryšys su atitinkamomis nukleininėmis bazėmis Atkreipkite dėmesį, kad RNR grandinė turi specifinę kryptį, kurią rodo 5 ir 3 galų fosfato liekanos.
4. Kadangi RNR molekulė yra viena grandinė, papildanti tik vieną iš DNR grandinių, guanino kiekis joje nebūtinai yra lygus citozino kiekiui, o adenino kiekis nebūtinai lygus uracilo kiekiui.
5. RNR gali būti hidrolizuojama šarminiais mononukleotidų 2,3-cikliniais diesteriais; 2, Y, 5- triesteris veikia kaip tarpinis hidrolizės produktas, kuris nesusidaro šarminės DNR hidrolizės metu, nes pastarojoje nėra 2-hidroksilo grupių; šarminis RNR labilumas (palyginti su DNR) yra naudinga savybė tiek diagnostikos, tiek analizės tikslais.
Informacija, esanti viengrandėje RNR, realizuojama kaip specifinė polimero grandinės purino ir pirimidino bazių seka (ty pirminėje struktūroje). Ši seka papildo koduojančią geno grandinę, iš kurios „skaitoma“ RNR. Dėl komplementarumo RNR molekulė gali specifiškai susijungti (hibridizuotis) su koduojančia grandine, bet nehibridizuotis su nekoduojančia DNR grandine. RNR seka (išskyrus T pakeitimą U) yra identiška nekoduojančios geno grandinės sekai (37.8 pav.).
Biologinės RNR funkcijos
Yra žinomi keli RNR tipai. Beveik visi jie tiesiogiai dalyvauja baltymų biosintezės procese. Citoplazminės RNR molekulės, kurios veikia kaip baltymų sintezės šablonai, vadinamos pasiuntinio RNR (mRNR). Kitas citoplazminės RNR tipas, ribosominė RNR (rRNR), atlieka struktūrinių ribosomų komponentų (organelių, vaidinančių svarbų vaidmenį baltymų sintezėje) vaidmenį. Transfer RNR (tRNR) adapterio molekulės dalyvauja iRNR informacijos vertime (vertime) į aminorūgščių seką baltymuose.
Didelė dalis RNR pirminių transkriptų, pagamintų eukariotinėse ląstelėse, įskaitant žinduolių ląsteles, suyra branduolyje ir nevaidina jokio struktūrinio ar informacinio vaidmens citoplazmoje. Išaugintame

Ryžiai. 37.7. Antrinė „kilpa su stiebu“ („plaukų segtukas“) tipo RNR molekulės struktūra, atsirandanti dėl intramolekulinio vandenilinių jungčių susidarymo tarp komplementarių nukleino bazių porų.
Žmogaus ląstelėse buvo aptikta mažų branduolinių RNR, kurios tiesiogiai nedalyvauja baltymų sintezėje, tačiau gali turėti įtakos RNR apdorojimui ir bendrai ląstelės „architektūrai“. Šių palyginti mažų molekulių dydžiai skiriasi, pastarosiose yra nuo 90 iki 300 nukleotidų (37.3 lentelė).
RNR yra pagrindinė kai kurių gyvūnų ir augalų virusų genetinė medžiaga. Kai kurie RNR virusai niekada nevyksta atvirkštinės RNR transkripcijos į DNR. Tačiau daugumai žinomų gyvūnų virusų, tokių kaip retrovirusai, būdinga atvirkštinė jų RNR genomo transkripcija, nukreipta nuo RNR priklausomos DNR polimerazės (atvirkštinės transkriptazės), kad susidarytų dvigrandė DNR kopija. Daugeliu atvejų gautas dvigrandės DNR nuorašas integruojamas į genomą ir toliau užtikrina viruso genų ekspresiją, taip pat naujų viruso RNR genomų kopijų gamybą.
RNR struktūrinė organizacija
Visuose eukariotuose ir prokariotiniuose organizmuose yra trys pagrindinės RNR molekulių klasės: informacinė (matrica arba pasiuntinys) RNR (mRNR), transportinė (tRNR) ir ribosominė (rRNR). Šių klasių atstovai skiriasi vienas nuo kito dydžiu, funkcijomis ir stabilumu.
Informacinė (mRNR) yra pati nevienalyčiausia klasė pagal dydį ir stabilumą. Visi šios klasės atstovai yra informacijos nešėjai iš geno į ląstelės baltymus sintezuojančią sistemą. Jie veikia kaip sintezuojamo polipeptido šablonai, tai yra nustato baltymo aminorūgščių seką (37.9 pav.).
Messenger RNR, ypač eukariotinės, turi keletą unikalių struktūrinių savybių. 5-asis mRNR galas yra „uždengtas“ 7-metilguanozintrifosfatu, prijungtu prie gretimo 2-0-metilribonukleozido 5-hidroksilo per trifosfato liekaną (37.10 pav.). mRNR molekulėse dažnai yra vidinių 6-metiladenino liekanų ir 2-0-metilintų ribonukleotidų. Nors „dangtelio“ reikšmė dar nėra iki galo išaiškinta, galima daryti prielaidą, kad gauta mRNR 5-galo struktūra naudojama specifiniam atpažinimui vertimo sistemoje. Baltymų sintezė prasideda nuo 5" (uždengto) mRNR galo. Kitame daugumos mRNR molekulių gale (3 gale) yra 20-250 nukleotidų poliadenilato grandinė. Konkrečios jos funkcijos nėra galutinai nustatytos. daryti prielaidą, kad ši struktūra yra atsakinga už intraląstelinio stabilumo mRNR palaikymą. Kai kurios mRNR, įskaitant histonines, neturi poli(A). Poli(A) buvimas mRNR struktūroje naudojamas atskirti nuo kitų RNR tipų frakcionuojant bendrą RNR. RNR ant kolonėlių su oligo(T), imobilizuota ant kieto pagrindo, pvz., celiuliozės, su kolonele atsiranda dėl papildomos poli(A) - "uodegos" sąveikos su imobilizuotu oligo (T).
Ryžiai. 37.8. Geno seka ir jo RNR nuorašas. Rodomos koduojančios ir nekoduojančios gijos, pažymimi jų poliškumas. RNR nuorašas, turintis poliškumą, yra papildantis koduojančią grandinę (su poliškumu 3–5) ir identiška savo seka (išskyrus T į U pakaitalus) ir nekoduojančios DNR grandinės poliškumu.

Ryžiai. 37.9. DNR genetinės informacijos ekspresija mRNR nuorašo forma ir vėlesnis vertimas dalyvaujant ribosomoms, kad susidarytų specifinė baltymo molekulė.
(žr. nuskaitymą)
Ryžiai. 37.10. Daugumos eukariotų pasiuntinių RNR 7-metilguanozintrifosfato 5 galuose esanti „dangtelio“ struktūra yra prijungta prie mRNR 5 galo. kuriame paprastai yra 2-O-metilpurino nukleotidas.
Žinduolių ląstelėse, įskaitant žmogaus ląsteles, subrendusios mRNR molekulės, esančios citoplazmoje, nėra visa transkribuotos geno srities kopija. Poliribonukleotidas, susidaręs dėl transkripcijos, yra citoplazminės mRNR pirmtakas; prieš paliekant branduolį, jis yra specialiai apdorojamas. Neapdoroti transkripcijos produktai, randami žinduolių ląstelių branduoliuose, sudaro ketvirtąją RNR molekulių klasę. Tokios branduolinės RNR yra labai nevienalytės ir pasiekia nemažus dydžius. Heterogeninės branduolinės RNR molekulių molekulinė masė gali būti didesnė nei , o iRNR molekulinė masė paprastai neviršija 2106. Jos apdorojamos branduolyje, o susidariusios subrendusios mRNR patenka į citoplazmą, kur tarnauja kaip baltymų matrica. biosintezė.
Pernešančios RNR (tRNR) molekulės paprastai turi apie 75 nukleotidus. Tokių molekulių molekulinė masė yra . tRNR susidaro ir dėl specifinio atitinkamų pirmtakų molekulių apdorojimo (žr. 39 skyrių). Transporto tRNR veikia kaip tarpininkai mRNR transliacijos metu. Bet kurioje ląstelėje yra mažiausiai 20 tipų tRNR molekulių. Kiekvienas tRNR tipas (kartais kelios rūšys) atitinka vieną iš 20 aminorūgščių, reikalingų baltymų sintezei. Nors kiekviena specifinė tRNR skiriasi nuo kitų savo nukleotidų seka, jos visos turi bendrų bruožų. Dėl kelių intrastrandą papildančių sričių visos tRNR turi antrinę struktūrą, vadinamą „dobilo lapeliu“ (37.11 pav.).
Visų tipų tRNR molekulės turi keturias pagrindines šakas. Akceptoriaus šaka susideda iš suporuotų nukleotidų „stiebo“ ir baigiasi CCA seka.Būtent per adenozilo liekanos Y-hidroksilo grupę įvyksta prisijungimas prie aminorūgšties karboksilo grupės. Likusios rankos taip pat susideda iš "stiebelių", sudarytų iš vienas kitą papildančių bazių porų ir nesuporuotų pagrindų kilpų (37.7 pav.). Antikodono grupė mRNR atpažįsta nukleotido tripletą arba kodoną (žr. 40 skyrių). D-šakis taip pavadintas dėl joje esančio dihidrouridino, o -šakis pavadintas pagal T-pseudouridino-C seką. Papildoma ranka yra pati kintamiausia struktūra ir naudojama kaip tRNR klasifikavimo pagrindas. 1 klasės tRNR (75 % viso jų skaičiaus) turi papildomą 3-5 bazinių porų ilgio ranką. Papildoma 2 klasės tRNR molekulių atšaka yra 13-21 bazinės poros ilgio ir dažnai apima nesuporuotą kilpą.

Ryžiai. 37.11. Aminoacil-tRNR molekulės, prie kurios 3-CCA-galo yra prijungta aminorūgštis, struktūra. Nurodomi intramolekuliniai vandenilio ryšiai ir antikodono, TTC ir dihidrouracilo gelių vieta. (Iš J. D. Watson. Molecular biology of the Gene 3rd, red.. Autorių teisės 1976, 1970, 1965, W. A. Benjamin, Inc., Menlo Park Calif.)
Antrinė struktūra, nulemta atitinkamų šakų nukleotidų bazių komplementarios sąveikos sistemos, būdinga visoms rūšims.Akceptorinėje šakoje yra septynios bazių poros, - rankoje - penkios bazių poros, D rankoje - trys (arba keturios) bazinės poros.
tRNR molekulės yra labai stabilios prokariotuose ir šiek tiek mažiau stabilios eukariotuose. Atvirkštinė situacija būdinga mRNR, kuri prokariotuose yra gana nestabili, o eukariotų organizmuose ji pasižymi dideliu stabilumu.
Ribosominė RNR. Ribosoma yra citoplazminė nukleoproteino struktūra, skirta baltymų sintezei iš mRNR šablono. Ribosoma suteikia specifinį kontaktą, dėl kurio įvyksta nukleotidų sekos, nuskaitytos iš konkretaus geno, vertimas į atitinkamo baltymo aminorūgščių seką.
Lentelėje. 37.2 pavaizduoti žinduolių ribosomų komponentai, kurių molekulinė masė yra 4,210 6 ir nusėdimo greitis (Swedberg vienetai). Žinduolių ribosomos susideda iš dviejų nukleoproteino subvienetų, didelių c
37.2 lentelė. Žinduolių ribosomų komponentai

molekulinė masė (60S) ir maža, kurios molekulinė masė (40S). 608 subvienetą sudaro 58-ribosomų RNR (rRNR), 5,8S-pRNR ir 28S-pRNR, taip pat daugiau nei 50 skirtingų polipeptidų. Mažas, 408 subvienetas apima vieną 18S-pRNR ir apie 30 polipeptidinių grandinių. Visos ribosominės RNR, išskyrus 5S-RNR, turi bendrą pirmtaką – 45S-RNR, esantį branduolyje (žr. 40 skyrių). 5S-RNR molekulė turi savo pirmtaką. Branduolėje labai metilintos ribosominės RNR yra supakuotos su ribosominiais baltymais. Citoplazmoje ribosomos yra gana stabilios ir gali atlikti daugybę vertimo ciklų.
Maža stabili RNR. Eukariotinėse ląstelėse rasta daug atskirų, labai konservuotų, mažų ir stabilių RNR molekulių. Dauguma šio tipo RNR yra ribonukleoproteinuose ir yra lokalizuotos branduolyje, citoplazmoje arba vienu metu abiejuose skyriuose. Šių molekulių dydžiai svyruoja nuo 90 iki 300 nukleotidų, jų kiekis yra 100 000-1 000 000 kopijų vienoje ląstelėje.
Mažos branduolinės ribonukleino dalelės (dažnai vadinamos snurps – iš anglų kalbos mažos branduolinės ribonukleino dalelės) tikriausiai vaidina esminį vaidmenį reguliuojant genų ekspresiją. Atrodo, kad U7 tipo nukleoproteinų dalelės dalyvauja formuojant 3-jų histono mRNR galų. Dalelės tikriausiai reikalingos poliadenilinimui, a intronų pašalinimui ir mRNR apdorojimui (žr. 39 skyrių). Skirtukas. 37.3. apibendrina kai kurias mažų stabilių RNR charakteristikas.
37.3 lentelė. Kai kurios mažos stabilios RNR rūšys randamos žinduolių ląstelėse

LITERATŪRA
Darnell J. ir kt. „Molecular Cell Biology“, „Scientific American Books“, 1986 m.
Hunt T. DNA Makes RNA Makes Protein, Elsevier, 1983. Lewin B. Genes, 2. leidimas, Wiley, 1985 m.
Turtingas A. ir kt. Kairiarankių Z-DNR chemija ir biologija, Annu. Rev. Biochem., 1984, 53, 847.
Turner P. Kontroliuojantys snurpso vaidmenys, Nature 1985, 316, 105. Watson J. D. The Double Helix, Atheneum, 1968.
Watsonas J.D., Crickas F.H.C. Nukleino rūgščių molekulinė struktūra. Gamta, 1953, 171, 737.
Zieve G. W. Dvi mažų stabilių RNR grupės, Cell, 1981, 25, 296.
Biologijos mokslų kandidatas S. GRIGOROVICH.
Ankstyviausioje savo istorijos aušroje, kai žmogus įgijo protą, o kartu ir gebėjimą mąstyti abstrakčiai, jis tapo nenugalimo poreikio viską paaiškinti kaliniu. Kodėl šviečia saulė ir mėnulis? Kodėl teka upės? Kaip pasaulis? Žinoma, vienas svarbiausių buvo gyvųjų esmės klausimas. Ryškus skirtumas tarp gyvų, augančių ir mirusių, nejudančių, buvo pernelyg ryškus, kad būtų galima ignoruoti.
Pirmasis virusas, kurį D. Ivanovskis aprašė 1892 m., yra tabako mozaikos virusas. Šio atradimo dėka tapo aišku, kad yra gyvų būtybių, primityvesnių už ląstelę.
Rusų mikrobiologas D.I.Ivanovskis (1864-1920), virusologijos įkūrėjas.
1924 metais A. I. Oparinas (1894-1980) pasiūlė, kad jaunos Žemės atmosferoje, kurią sudarė vandenilis, metanas, amoniakas, anglies dioksidas ir vandens garai, galima susintetinti aminorūgštis, kurios vėliau spontaniškai susijungė į baltymus.
Amerikiečių biologas Oswaldas Avery, atlikdamas eksperimentus su bakterijomis, įtikinamai įrodė, kad būtent nukleino rūgštys yra atsakingos už paveldimų savybių perdavimą.
Lyginamoji RNR ir DNR struktūra.
Paprasčiausio organizmo Tetrahymena ribozimo dvimatė erdvinė struktūra.
Scheminis ribosomos, baltymų sintezės molekulinės mašinos, vaizdas.
„Evoliucijos in vitro“ proceso schema (Selekso metodas).
Louis Pasteur (1822-1895) pirmasis atrado, kad tos pačios medžiagos – vyno rūgšties – kristalai gali turėti dvi veidrodiškai simetriškas erdvines konfigūracijas.
Penktojo dešimtmečio pradžioje Stanley Milleris iš Čikagos universiteto (JAV) atliko pirmąjį eksperimentą, kuris imitavo chemines reakcijas, kurios gali vykti jaunos Žemės sąlygomis.
Chiralinės molekulės, tokios kaip aminorūgštys, yra veidrodiškai simetriškos, kaip ir kairiosios bei dešinės rankos. Pats terminas „chiralumas“ kilęs iš graikiško žodžio „chiros“ – ranka.
RNR pasaulio teorija.
Mokslas ir gyvenimas // Iliustracijos
Kiekvienu istorijos etapu žmonės siūlydavo savo sprendimą dėl gyvybės atsiradimo mūsų planetoje mįslės. Senoliai, nemokėję žodžio „mokslas“, rado paprastą ir prieinamą nežinomybės paaiškinimą: „Viskas, kas yra aplinkui, kažkada buvo kažkada sukurta. Taip atsirado dievai.
Nuo senovės civilizacijų gimimo Egipte, Kinijoje, o vėliau ir šiuolaikinio mokslo lopšyje – Graikijoje, iki viduramžių „autoritetų“ pastebėjimai ir nuomonės buvo pagrindinis pasaulio pažinimo metodas. Nuolatiniai stebėjimai vienareikšmiškai liudijo, kad gyvieji, esant tam tikroms sąlygoms, atsiranda iš negyvų: uodai ir krokodilai – iš pelkių purvo, musės – iš pūvančio maisto, o pelės – iš nešvarių skalbinių, apibarstytų kviečiais. Svarbu tik laikytis tam tikros temperatūros ir drėgmės.

Viduramžių Europos „mokslininkai“, remdamiesi religine pasaulio sukūrimo dogma ir dieviškų planų nesuvokiamumu, ginčytis dėl gyvybės kilmės manė galima tik Biblijos ir religinių raštų rėmuose. To, ką Dievas sukūrė, esmė negali būti suvokiama, o gali būti tik „nurodyta“ naudojant informaciją iš šventų tekstų arba esant dieviškojo įkvėpimo įtakai. Hipotezių tikrinimas tuo metu buvo laikomas blogu manieru, o bet koks bandymas suabejoti šventosios bažnyčios nuomone – nemalonus poelgis, erezija ir šventvagystė.
Žinios apie gyvenimą žengė į vandenį. Senovės Graikijos filosofų pasiekimai išliko mokslinės minties viršūne du tūkstančius metų. Reikšmingiausi iš jų buvo Platonas (428/427 – 347 m. pr. Kr.) ir jo mokinys Aristotelis (384 – 322 m. pr. Kr.). Platonas, be kita ko, pasiūlė idėją pagyvinti iš pradžių negyvą materiją, nes į ją įsilieja nemirtinga nemateriali siela - „psichika“. Taip atsirado teorija apie spontanišką gyvų būtybių susidarymą iš negyvų dalykų.
Didysis mokslo žodis „eksperimentas“ atsirado Renesanso laikais. Prireikė dviejų tūkstančių metų, kad žmogus nuspręstų suabejoti autoritetingų senovės mokslininkų teiginių nekintamumu. Vienas pirmųjų mums žinomų drąsuolių buvo italų gydytojas Francisco Redi (1626–1698). Jis atliko itin paprastą, bet veiksmingą eksperimentą: įdėjo mėsos gabalą į kelis indus, vieną iš jų uždengė tankiu audiniu, kitus – marle, o trečią paliko atvirą. Tai, kad musių lervos vystėsi tik atviruose induose (kuriuose galėjo nutūpti musės), bet ne uždaruose (kurie vis dar turėjo prieigą prie oro), smarkiai prieštaravo Platono ir Aristotelio šalininkų įsitikinimams apie pro šalį besiveržiančią nesuvokiamą gyvybės jėgą. orą ir negyvą materiją paverčiant gyva medžiaga.
Šiuo ir panašiais eksperimentais prasidėjo įnirtingų kovų tarp dviejų mokslininkų grupių – vitalistų ir mechanikų – laikotarpis. Ginčo esmė buvo klausimas: „Ar gyvų būtybių funkcionavimą (ir išvaizdą) galima paaiškinti fiziniais dėsniais, kurie taikomi ir negyvajai medžiagai? Vitalistai jam atsakė neigiamai. „Ląstelė – tik iš ląstelės, visa gyva – tik iš gyvos! Ši pozicija, iškelta XIX amžiaus viduryje, tapo vitalizmo vėliava. Paradoksalu šiame ginče yra tai, kad net ir šiandien, žinodami apie mūsų kūną sudarančių atomų ir molekulių „negyvąją“ prigimtį ir iš esmės sutikdami su mechaniniu požiūriu, mokslininkai neturi eksperimentinio kilmės galimybės patvirtinimo. ląstelių gyvybės iš negyvos medžiagos. Dar niekam nepavyko „sudaryti“ net pačios primityviausios ląstelės iš „neorganinių“ „detalių“, esančių už gyvų organizmų ribų. Taigi, galutinis taškas šiame epochiniame ginče dar turi būti nustatytas.
Taigi, kaip Žemėje galėjo atsirasti gyvybė? Dalijantis mechanizatorių pozicijomis, tikrai lengviausia įsivaizduoti, kad gyvybė pirmiausia turėjo atsirasti kažkokia labai paprasta, primityviai sutvarkyta forma. Tačiau, nepaisant struktūros paprastumo, tai vis tiek turi būti Gyvenimas, tai yra kažkas, kas turi minimalų savybių rinkinį, skiriantį gyvą nuo negyvojo.
Kas tai yra, šios gyvybiškai svarbios savybės? Kas iš tikrųjų skiria gyvuosius nuo negyvų?
Iki XIX amžiaus pabaigos mokslininkai buvo įsitikinę, kad viskas, kas gyva, yra sukurta iš ląstelių, ir tai yra ryškiausias skirtumas tarp jos ir negyvos materijos. Apie tai buvo galvota dar prieš atrandant virusus, kurie, nors ir mažesni už visas žinomas ląsteles, gali aktyviai užkrėsti kitus organizmus, daugintis juose ir susilaukti palikuonių su tokiomis pat (arba labai panašiomis) biologinėmis savybėmis. Pirmąjį atrastą virusą – tabako mozaikos virusą – aprašė rusų mokslininkas Dmitrijus Ivanovskis (1864-1920) 1892 m. Nuo tada tapo aišku, kad būtybės, primityvesnės už ląsteles, taip pat gali pretenduoti į teisę būti vadinamos Gyvybe.
Virusų, o vėliau ir dar primityvesnių gyvų būtybių formų – viroidų atradimas ilgainiui leido suformuluoti minimalų savybių rinkinį, kuris yra būtinas ir pakankamas, kad tiriamas objektas būtų vadinamas gyvu. Pirma, jis turi sugebėti atgaminti savo rūšį. Tačiau tai nėra vienintelė sąlyga. Jei hipotetinė pirmykštė gyvybės substancija (pavyzdžiui, primityvi ląstelė ar molekulė) galėtų tiesiog sukurti tikslias savo kopijas, ji galiausiai negalėtų išgyventi kintančiomis aplinkos sąlygomis jaunoje Žemėje ir formuojantis kitiems. , sudėtingesnės formos (evoliucija) taptų neįmanomos. Todėl mūsų tariamą primityvią „pirmojo gyvenimo substanciją“ galima apibrėžti kaip tai, kas sutvarkyta kuo paprasčiau, bet tuo pačiu galinti keisti ir perduoti savo savybes palikuonims.
RNR, kaip taisyklė, susideda iš vienos grandinės, susuktos į spiralę. Virusai turi dvigrandę RNR. RNR randama branduolyje, branduolyje, citoplazmoje, ribosomose. RNR molekulės yra trumpesnės nei DNR molekulės.
RNR tipai
Yra trys RNR tipai: ribosominė, pasiuntinio (mRNR) ir transportavimo (tRNR). Jie skiriasi vienas nuo kito pagal vietą ląstelėje, dydį, nukleotidų sudėtį ir funkcines savybes.
RNR, kurią sintetina fermentai RNR polimerazės ant DNR molekulės. DNR molekulės sekcijos nukleotidų seka lemia nukleotidų išsidėstymo tvarką RNR molekulėje.
Daugumoje ląstelių RNR kiekis yra daug didesnis (nuo 5 iki 10 kartų) nei DNR. Didžioji RNR dalis yra ribosominė.
RNR funkcijos
RNR funkcijos: įgyvendina paveldimą informaciją, dalyvauja baltymų sintezėje.
Informacinis(matrica) RNR (mRNR) yra DNR dalies, ty vieno ar kelių genų, kopija. Jis perduoda genetinę informaciją į polipeptidinės grandinės sintezės vietą ir tiesiogiai joje dalyvauja. Pagal DNR sekcijos ilgį, kurią kopijuoja RNR, ji susideda iš 300-30 000 nukleotidų. Dalis ir RNR ląstelėje sudaro apie 5% viso. Molekulės ir RNR yra gana nestabilios – greitai skyla į nukleotidus. Jų gyvenimo trukmė eukariotinėse ląstelėse siekia iki kelių valandų, mikroorganizmuose – kelias minutes.
Kaip ir DNR molekulė, RNR taip pat turi antrines ir tretines struktūras, kurios susidaro naudojant vandenilinius ryšius, hidrofobines, elektrostatines sąveikas ir kt.
Ribosominis RNR sudaro 60% ribosomų masės, apie 85% viso RNR kiekio ląstelėje. Apima 3000-5000 nukleotidų. Ji nedalyvauja perduodant paveldimą informaciją. Ji yra ribosomos dalis ir sąveikauja su jos baltymais, kurių eukariotuose yra apie 100. Eukariotai turi keturių tipų ribosomų RNR, o prokariotai – tris. Atlieka struktūrinę funkciją: suteikia tam tikrą erdvinį mRNR ir tRNR išsidėstymą ribosomoje.
Transportas (tRNR) – perneša aminorūgštis į baltymų sintezės vietą. Pagal komplementarumo principą jis atpažįsta mRNR sritį, atitinkančią pernešamą aminorūgštį. Kiekviena aminorūgštis yra pernešama į baltymų sintezės vietą savo tRNR. tRNR transportuoja ląstelės citoskeleto elementai.
Jis turi trilapio (dobilo lapo) formą – nuolatinę antrinę struktūrą, kurią užtikrina vandeniliniai ryšiai. tRNR viršuje yra nukleotidų tripletas, atitinkantis mRNR kodoną ir vadinamas antikodonas . Netoli bazės yra vieta, prie kurios dėl kovalentinio ryšio yra prijungta aminorūgšties molekulė. Sudėtyje yra 70-90 nukleotidų tRNR. Sudaro iki 10% viso RNR kiekio. Yra žinoma apie 60 tRNR tipų.
tRNR gali turėti gana kompaktišką L formos netaisyklingą formą tretinis struktūra.
Dinukleotidai
Jie susideda iš dviejų nukleotidų, tačiau turi struktūrinių savybių. Garsiausi yra: nikotinamido adenino dinukleotidas (NAD +), nikotinamido adenino dinukleotido fosfatas (NADP +). Pagrindinė funkcija yra elektronų (2) ir vandenilio jonų (1) perdavimas. Gali atsigauti:
VIRŠ + + 2e - + H + → NADH;
NADP + + 2e - + H + → NADPH.
Tam tikroje kai kurių reakcijų vietoje šie junginiai dovanoja vandenilio protoną, elektronus:
NADH → VIRŠ + + 2e - + H + ;
NADPH → NADP + + 2e - + H +
Priklausomai nuo to, kuris monosacharidas yra polinukleotido struktūriniame vienete - ribozė arba 2-dezoksiribozė, išskirti
- ribonukleino rūgštys(RNR) ir
- dezoksiribonukleino rūgštys(DNR).
DNR makromolekulių nukleotidų vienetuose gali būti adeninas, guaninas, citozinas ir timinas. RNR sudėtis skiriasi tuo, kad timinas pateikti uracilas.
DNR molekulinė masė siekia dešimtis milijonų amu. Tai yra ilgiausios žinomos makromolekulės. RNR molekulinė masė yra daug mažesnė (nuo kelių šimtų iki dešimčių tūkstančių). DNR daugiausia randama ląstelių branduoliuose, RNR – ląstelių ribosomose ir protoplazmoje.
Apibūdinant nukleorūgščių struktūrą, atsižvelgiama į skirtingus makromolekulių organizavimo lygius: pirminis ir antraeilis struktūra.
- Pirminė struktūra Nukleorūgštys yra nukleotidų sudėtis ir tam tikra nukleotidų seka polimero grandinėje.
Sutrumpintame vienos raidės žymėjime ši struktūra rašoma kaip ...– A – G – C –...
- Pagal antrinė struktūra nukleorūgštys supranta erdviškai išdėstytas polinukleotidinių grandinių formas.

Šią trimatę struktūrą palaiko daugybė vandenilio jungčių, sudarytų iš azoto bazių, nukreiptų į spiralės vidų. Vandenilinės jungtys susidaro tarp vienos grandinės purino bazės ir kitos grandinės pirimidino bazės. Šios bazės sudaro viena kitą papildančias poras (iš lat. komplementum- papildymas). Vandeniliniai ryšiai tarp papildomų bazių porų susidaro dėl jų erdvinio atitikimo. Pirimidino bazė papildo purino bazę:

Vandeniliniai ryšiai tarp kitų bazių porų neleidžia joms tilpti į dvigubos spiralės struktūrą. Šiuo būdu,
- TIMINAS (T) papildo adeniną (A),
- Citozinas (C) papildo GUANINĄ (G).

Polinukleotidinių grandinių komplementarumas yra pagrindinės DNR saugojimo ir paveldimų požymių perdavimo funkcijos cheminis pagrindas.
DNR gebėjimą ne tik saugoti, bet ir panaudoti genetinę informaciją lemia šios jos savybės:
Antrinė RNR struktūra. Skirtingai nuo DNR, RNR molekulės susideda iš vienos polinukleotidinės grandinės ir neturi griežtai apibrėžtos erdvinės formos (antrinė RNR struktūra priklauso nuo jų biologinių funkcijų).
Pagrindinis RNR vaidmuo yra tiesioginis dalyvavimas baltymų biosintezėje. Yra žinomi trys ląstelių RNR tipai, kurie skiriasi savo vieta ląstelėje, sudėtimi, dydžiu ir savybėmis, kurios lemia specifinį jų vaidmenį formuojant baltymų makromolekules:
- informacinės (matricos) RNR perduoda DNR užkoduotą informaciją apie baltymo struktūrą iš ląstelės branduolio į ribosomas, kuriose vyksta baltymų sintezė;
- pernešančios RNR surenka aminorūgštis ląstelės citoplazmoje ir perneša jas į ribosomą; Šio tipo RNR molekulės iš atitinkamų pasiuntinio RNR grandinės atkarpų „sumoka“, kurios aminorūgštys turi dalyvauti baltymų sintezėje;
- ribosominė RNR užtikrina tam tikros struktūros baltymų sintezę, nuskaito informaciją iš informacinės (matricos) RNR.



