A plasztidok szerkezete: 1 - külső membrán; 2 - belső membrán; 3 - stroma; 4 - tilakoid; 5 - gabona; 6 - lamellák; 7 - keményítőszemek; 8 - lipidcseppek.
A plasztidok csak a növényi sejtekben találhatók. Megkülönböztetni a plasztidok három fő típusa: leukoplasztok - színtelen plasztiszok a festetlen növényrészek sejtjeiben, kromoplasztok - színes plasztiszok általában sárga, vörös és narancssárga, kloroplasztok - zöld plasztiszok.
Kloroplasztok. A magasabb rendű növények sejtjeiben a kloroplasztiszok bikonvex lencse alakúak. A kloroplasztiszok hossza 5-10 mikron, átmérője 2-4 mikron. A kloroplasztokat két membrán határolja. A külső membrán (1) sima, a belső (2) összetett hajtogatott szerkezetű. A legkisebb redőt ún tilakoid(4). A tilakoidok egy csoportját, amelyek úgy vannak egymásra rakva, mint egy köteg érme, az úgynevezett csiszolt(5). A kloroplaszt átlagosan 40-60, sakktáblás mintázatú szemcsét tartalmaz. A granulátumok lapított csatornákkal kapcsolódnak egymáshoz - lamellák(6). A tilakoid membránok fotoszintetikus pigmenteket és enzimeket tartalmaznak, amelyek biztosítják az ATP szintézist. A fő fotoszintetikus pigment a klorofill, amely meghatározza a kloroplasztiszok zöld színét.
A kloroplasztiszok belső tere ki van töltve stroma(3). A stroma kör alakú csupasz DNS-t, 70S típusú riboszómákat, Calvin-ciklus enzimeket és keményítőszemcséket tartalmaz (7). Minden tilakoid belsejében protontároló található, a H + felhalmozódik. A kloroplasztiszok a mitokondriumokhoz hasonlóan kettészakadva képesek autonóm szaporodásra. A magasabb rendű növények zöld részeinek sejtjeiben találhatók, különösen sok kloroplasztisz a levelekben és a zöld gyümölcsökben. Az alacsonyabb rendű növények kloroplasztjait kromatoforoknak nevezzük.
A kloroplasztok funkciói: fotoszintézis. Úgy gondolják, hogy a kloroplasztiszok ősi endoszimbiotikus cianobaktériumokból származnak (szimbiogenezis elmélet). Ennek a feltételezésnek az alapja a kloroplasztiszok és a modern baktériumok sokféle hasonlósága (kör alakú, "csupasz" DNS, 70S típusú riboszómák, szaporodási mód).
Leukoplasztok. A forma változó (gömb alakú, lekerekített, köpölyözött stb.). A leukoplasztokat két membrán határolja. A külső membrán sima, a belső kis tilakoidokat képez. A stroma körkörös "csupasz" DNS-t, 70S típusú riboszómákat, tartalék tápanyagok szintéziséhez és hidrolíziséhez szükséges enzimeket tartalmaz. Nincsenek pigmentek. Különösen sok leukoplasztban találhatók a növény földalatti szerveinek sejtjei (gyökerek, gumók, rizómák stb.). A leukoplasztok funkciói: tartalék tápanyagok szintézise, felhalmozódása és tárolása. Amiloplasztok- keményítőt szintetizáló és felhalmozó leukoplasztok, elaioplasztok- olajok, proteinoplasztok- fehérjék. Ugyanabban a leukoplasztban különböző anyagok halmozódhatnak fel.
Kromoplasztok. Két membrán korlátozza. A külső membrán sima, a belső vagy szintén sima, vagy egyedi tilakoidokat alkot. A sztróma körkörös DNS-t és pigmenteket - karotinoidokat - tartalmaz, amelyek sárga, piros vagy narancssárga színt adnak a kromoplasztoknak. A pigmentek felhalmozódásának formája eltérő: kristályok formájában, lipidcseppekben oldva (8), stb. Az érett gyümölcsök, szirmok, őszi levelek sejtjeiben, ritkán gyökérnövények sejtjeiben található. A kromoplasztokat a plasztiszok fejlődésének utolsó szakaszának tekintik.
A kromoplasztok funkciói: a virágok és a gyümölcsök színezése, és ezáltal vonzza a beporzókat és a magvak szétszóródását.
Proplasztidokból mindenféle plasztisz előállítható. proplasztidok- a merisztémás szövetekben található kis organellumok. Mivel a plasztidok közös eredetûek, lehetségesek közöttük az interkonverziók. A leukoplasztok kloroplasztokká alakulhatnak (a burgonyagumók zöldítése a fényben), a kloroplasztiszok - kromoplasztokká (a levelek sárgulása és a gyümölcsök pirosodása). A kromoplasztok leukoplasztokká vagy kloroplasztokká történő átalakulását lehetetlennek tartják.
A plasztidák az autotróf növényi sejtek fő citoplazmatikus szervei. A név a görög "plastos" szóból származik, ami azt jelenti, hogy "faragott".
A plasztidok fő funkciója a szerves anyagok szintézise, a saját DNS és RNS jelenléte, valamint a fehérjeszintézis szerkezete miatt. A plasztidok pigmenteket is tartalmaznak, amelyek meghatározzák a színüket. Ezeknek az organellumoknak minden típusa összetett belső szerkezettel rendelkezik. Kívül a plasztidot két elemi membrán fedi, belső membránrendszer van, amely a stromába vagy mátrixba merül.
A plasztidok szín és funkció szerinti osztályozása magában foglalja ezeket az organellumokat három típusra osztva: kloroplasztokra, leukoplasztokra és kromoplasztokra. Az algák plasztidjait kromatoforoknak nevezzük.
Ezek magasabb rendű növények zöld plasztidjai, amelyek klorofillt, egy fotoszintetikus pigmentet tartalmaznak. Lekerekített testek, 4-10 mikron méretűek. A kloroplaszt kémiai összetétele: körülbelül 50% fehérje, 35% zsír, 7% pigmentek, kis mennyiségű DNS és RNS. A különböző növénycsoportok képviselőinél a színt meghatározó és a fotoszintézisben részt vevő pigmentek komplexe eltérő. Ezek a klorofill és a karotinoidok (xantofill és karotin) altípusai.
Fénymikroszkóp alatt szemlélve a plasztidok szemcsés szerkezete látható – ezek a grana. Elektronmikroszkóp alatt kis átlátszó, lapított tasakok (ciszterna vagy grana) figyelhetők meg, amelyeket fehérje-lipid membrán képez, és közvetlenül a stromában helyezkednek el. Sőt, némelyikük érmeoszlopoknak (thylakoid grana) hasonlító csomagokba csoportosul, mások, nagyobbak, a tilakoidok között helyezkednek el. Ennek a szerkezetnek köszönhetően megnő a grana lipid-fehérje-pigment komplexének aktív szintetizáló felülete, melyben fényben fotoszintézis megy végbe.
Ezek a plasztidok, amelyek színe sárga, narancssárga vagy vörös, a karotinoidok felhalmozódása miatt. A kromoplasztok jelenléte miatt az őszi levelek, virágszirmok, az érett gyümölcsök (paradicsom, alma) jellegzetes színt kapnak. Ezek az organellumok különböző formájúak lehetnek - kerekek, sokszögűek, néha tű alakúak.

Leukoplasztok
Színtelen plasztiszok, amelyek fő funkciója általában a tárolás. Ezek az organellumok viszonylag kicsik. Kerekek vagy enyhén hosszúkásak, minden élő növényi sejtre jellemzőek. A leukoplasztokban a szintézist bonyolultabbak egyszerű vegyületeiből - keményítőből, zsírokból, fehérjékből - hajtják végre, amelyeket gumókban, gyökerekben, magvakban, gyümölcsökben tárolnak. Elektronmikroszkóp alatt észrevehető, hogy minden leukoplasztot kétrétegű membrán borít, a stromában csak egy vagy kis számú membránkinövés található, a fő teret szerves anyagok töltik ki. Attól függően, hogy milyen anyagok halmozódnak fel a stromában, a leukoplasztokat amiloplasztokra, proteinoplasztokra és eleoplasztokra osztják.
Kloroplaszt
A plasztidok membránszervecskék, amelyek a fotoszintetikus eukarióta szervezetekben (magasabb növények, alacsonyabb szintű algák, egyes egysejtűek) találhatók. A plasztidokat két membrán veszi körül, mátrixuk saját genomiális rendszerrel rendelkezik, a plasztidok funkciói a sejt energiaellátásához kapcsolódnak, ami a fotoszintézis szükségleteihez megy.
A magasabb rendű növényekben különféle plasztidok egész halmazát találták (kloroplaszt, leukoplaszt, amiloplaszt, kromoplaszt), amelyek az egyik plasztisztípus kölcsönös átalakulásának sorozata a másikba. A fotoszintetikus folyamatokat végző fő szerkezet a kloroplaszt.
Magasabb növényekben az érett kloroplasztiszok osztódása is előfordul, de nagyon ritkán. A kloroplasztiszok számának növekedése és a plasztiszok egyéb formái (leukoplasztok és kromoplasztok) képződését a prekurzor struktúrák, proplasztidok átalakulásának útjaként kell tekinteni. A különféle plasztidok fejlődési folyamata egy monotróp (egy irányba haladó) alakváltozási sorozatként ábrázolható:
Számos tanulmány megállapította a plasztidok ontogenetikai átmeneteinek visszafordíthatatlan természetét. A magasabb rendű növényekben a kloroplasztiszok megjelenése és fejlődése a proplasztidok változásán keresztül történik. A proplasztidok kicsi (0,4-1 μm) kétmembrános vezikulák, amelyek belső szerkezetükben nincsenek megkülönböztető jegyek. A citoplazmatikus vakuoláktól sűrűbb tartalmukban és két, külső és belső határoló membrán jelenlétében különböznek. A belső membrán kis redőket vagy kis vakuolákat képezhet. A proplasztidok leggyakrabban az osztódó növényi szövetekben (gyökér merisztéma sejtjei, levelei, szárak növekedési pontjain stb.) találhatók. Számuk minden valószínűség szerint megnövekszik az osztódás vagy bimbózás, a kis kétmembrános hólyagok elválasztása a proplasztid testtől.
Kloroplasztok
 A kloroplasztok olyan szerkezetek, amelyekben fotoszintetikus folyamatok mennek végbe, ami végső soron szén-dioxid megkötéséhez, oxigén felszabadulásához és cukrok szintéziséhez vezet. 2-4 mikron szélességű és 5-10 mikron hosszúságú hosszúkás szerkezetek. A zöld algák óriási kloroplasztiszokkal (kromatoforokkal) rendelkeznek, amelyek hossza eléri az 50 mikront.
A kloroplasztok olyan szerkezetek, amelyekben fotoszintetikus folyamatok mennek végbe, ami végső soron szén-dioxid megkötéséhez, oxigén felszabadulásához és cukrok szintéziséhez vezet. 2-4 mikron szélességű és 5-10 mikron hosszúságú hosszúkás szerkezetek. A zöld algák óriási kloroplasztiszokkal (kromatoforokkal) rendelkeznek, amelyek hossza eléri az 50 mikront.
a zöld algák sejtenként egy kloroplasztot tartalmazhatnak. A magasabb rendű növények sejtjeiben általában átlagosan 10-30 kloroplaszt található. Vannak olyan sejtek, amelyekben hatalmas számú kloroplaszt található. Például körülbelül 1000 kloroplasztot találtak a bokor palánkszövetének óriás sejtjeiben.
A kloroplasztok olyan szerkezetek, amelyeket két membrán határol - belső és külső. A külső membrán, a belsőhöz hasonlóan, körülbelül 7 µm vastagságú, ezeket egy körülbelül 20-30 nm közötti membránköz választja el egymástól. A kloroplasztiszok belső membránja választja el a plasztid stromát, hasonlóan a mitokondriális mátrixhoz. A magasabb rendű növények érett kloroplasztiszának strómájában kétféle belső membrán látható. Ezek olyan membránok, amelyek lapos, kiterjesztett stromalamellákat és tilakoid membránokat, lapos korong alakú vakuolákat vagy tasakokat képeznek.
A stroma lamellák (körülbelül 20 μm vastagok) lapos üreges zsákok, vagy úgy néznek ki, mint egy elágazó és egymással összefüggő csatornák hálózata, amelyek ugyanabban a síkban helyezkednek el. Általában a kloroplasztisz belsejében lévő stroma lamellái egymással párhuzamosan fekszenek, és nem képeznek kapcsolatot egymással.
A kloroplasztiszokban a stroma membránjain kívül hártyás tilakoidok találhatók. Ezek lapos zárt membrántasakok, amelyek korong alakúak. A membránközi tér mérete is körülbelül 20-30 nm. Az ilyen tilakoidok érmeoszlophoz hasonló halmokat képeznek, amelyeket grana-nak neveznek.
A tilakoidok szemenkénti száma nagyon változó, néhánytól 50-ig vagy még többig. Az ilyen halmok mérete elérheti a 0,5 μm-t, így a szemcsék egyes tárgyakon fénymikroszkópban is láthatóak. A magasabb rendű növények kloroplasztiszában a szemek száma elérheti a 40-60-at is. A gránában lévő tilakoidok olyan közel vannak egymáshoz, hogy membránjaik külső rétegei szorosan kapcsolódnak egymáshoz; tilakoid membránok találkozásánál körülbelül 2 nm vastag sűrű réteg képződik. A gránában a tilakoidok zárt kamrái mellett általában lamellák szakaszai is vannak, amelyek membránjuk és tilakoid membránjuk érintkezési pontjain szintén sűrű, 2 nm-es rétegeket alkotnak. A stroma lamellák tehát úgy tűnik, hogy összekapcsolják a kloroplaszt egyes szemcséit. A tilakoid kamrák üregei azonban mindig zártak, és nem jutnak át a stroma lamellák intermembrán terének kamráiba. A stroma lamellák és a tilakoid membránok a plasztid fejlődésének kezdeti szakaszában a belső membrántól való elválasztással jönnek létre.
A kloroplasztiszok mátrixában (sztrómájában) DNS-molekulák és riboszómák találhatók; ott van a tartalék poliszacharid, a keményítő elsődleges lerakódása is keményítőszemcsék formájában.
A kloroplasztiszokra jellemző a pigmentek, klorofillok jelenléte, amelyek színt adnak a zöld növényeknek. A zöld növények a klorofill segítségével elnyelik a napfény energiáját és kémiai energiává alakítják.
A kloroplasztiszok funkciói
plasztid genom
A mitokondriumokhoz hasonlóan a kloroplasztiszoknak is megvan a maguk genetikai rendszere, amely számos fehérje szintézisét biztosítja magukon a plasztidokon belül. A kloroplasztiszok mátrixában DNS, különféle RNS és riboszómák találhatók. Kiderült, hogy a kloroplasztiszok DNS-e élesen eltér a sejtmag DNS-étől. Legfeljebb 40-60 mikron hosszúságú, 0,8-1,3x108 dalton molekulatömegű ciklikus molekulák képviselik. Egy kloroplasztban sok DNS-másolat lehet. Tehát egy kukorica kloroplasztiszban 20-40 DNS-molekula másolata van. A ciklus időtartama és a nukleáris és kloroplasztisz-DNS replikációs sebessége, amint azt a zöld algasejtek mutatják, nem egyezik. A kloroplaszt DNS nincs komplexben hisztonokkal. A kloroplaszt DNS-nek mindezek a tulajdonságai közel állnak a prokarióta sejt DNS-éhez. Sőt, a DNS hasonlóságát a kloroplasztiszok és a baktériumok között az is alátámasztja, hogy a fő transzkripciót szabályozó szekvenciák (promoterek, terminátorok) megegyeznek. Az RNS minden típusa (hírvivő, transzfer, riboszómális) szintetizálódik a kloroplasztiszok DNS-én. A kloroplaszt DNS az rRNS-t kódolja, amely ezen plasztidok riboszómáinak része, amelyek a prokarióta 70S típusba tartoznak (16S és 23S rRNS-t tartalmaznak). A kloroplaszt riboszómák érzékenyek a klóramfenikol nevű antibiotikumra, amely gátolja a prokarióta sejtekben a fehérjeszintézist.
Csakúgy, mint a kloroplasztiszok esetében, itt is egy speciális fehérjeszintézis rendszer létezésével állunk szemben, amely különbözik a sejtben lévőtől.
Ezek a felfedezések felkeltették az érdeklődést a kloroplasztiszok szimbiotikus eredetének elmélete iránt. A 19. és 20. század fordulóján megfogalmazott gondolat, hogy a kloroplasztiszok heterotróf sejtek és prokarióta kékalgák kombinálásával keletkeztek. (A.S. Fomintsin, K.S. Merezhkovsky) ismét megerősítést talál. Ezt az elméletet támasztja alá a kloroplasztiszok és a kék-zöld algák szerkezetének elképesztő hasonlósága, a hasonlóság fő funkcionális jellemzőikkel, és elsősorban a fotoszintetikus folyamatokra való képességükkel.
Számos tény ismeretes a kékalgák valódi endoszimbiózisáról alacsonyabb rendű növények és protozoonok sejtjeivel, ahol működnek és látják el a gazdasejtet fotoszintézis termékekkel. Kiderült, hogy az izolált kloroplasztokat egyes sejtek is szelektálhatják és endoszimbiontaként használhatják fel. Számos gerinctelen állatnál (forgófélék, puhatestűek), amelyek magasabb algákkal táplálkoznak, amelyeket megemésztenek, az ép kloroplasztok az emésztőmirigyek sejtjeiben vannak. Így egyes növényevő puhatestűek sejtjeiben ép kloroplasztiszokat találtak működőképes fotoszintetikus rendszerrel, amelyek aktivitását C14O2 beépülésével követték nyomon.
Mint kiderült, a kloroplasztok pinocitózissal juttathatók be az egér fibroblasztsejtek citoplazmájába. A hidrolázok azonban nem támadták meg őket. Az ilyen sejtek, amelyekben zöld kloroplasztok is voltak, öt generáción belül osztódhattak, miközben a kloroplasztiszok érintetlenek maradtak és fotoszintetikus reakciókat hajtottak végre. A kloroplasztiszokat mesterséges közegben próbálták tenyészteni: a kloroplasztiszok fotoszintetizálni tudtak, RNS-szintézis zajlott bennük, 100 órán át épek maradtak, osztódások még 24 órán belül is megfigyelhetők voltak. De aztán visszaesett a kloroplasztiszok aktivitása, és meghaltak.
Ezek a megfigyelések és számos biokémiai vizsgálat kimutatta, hogy a kloroplasztiszok autonómiájának jellemzői még mindig nem elegendőek funkcióik hosszú távú fenntartásához, és még inkább szaporodásukhoz.
A közelmúltban sikerült teljesen megfejteni a magasabb rendű növényi kloroplasztiszok ciklikus DNS-molekulájában található nukleotidok teljes szekvenciáját. Ez a DNS akár 120 gént is kódolhat, köztük: 4 riboszómális RNS génjeit, kloroplasztiszok 20 riboszomális fehérjéjét, kloroplasztisz RNS polimeráz egyes alegységeinek génjeit, az I és II fotorendszerek több fehérjét, az ATP szintetáz 12 alegységéből 9-et, részeket. elektrontranszport lánc komplexek fehérjéi, a ribulóz-difoszfát-karboxiláz egyik alegysége (a CO2-kötés kulcsenzime), 30 tRNS-molekula és további 40, még ismeretlen fehérje. Érdekes módon a kloroplasztiszok DNS-ében hasonló génkészletet találtak magasabb rendű növények olyan távoli képviselőiben, mint a dohány és a májmoha.
A kloroplaszt fehérjék fő tömegét a nukleáris genom szabályozza. Kiderült, hogy számos legfontosabb fehérje, enzim, és ennek megfelelően a kloroplasztiszok anyagcsere-folyamatai a sejtmag genetikai irányítása alatt állnak. Tehát a sejtmag szabályozza a klorofill, karotinoidok, lipidek, keményítő szintézisének egyes szakaszait. Számos sötét stádiumú enzim és más enzim nukleáris ellenőrzés alatt áll, beleértve az elektrontranszport lánc egyes összetevőit. A nukleáris gének a kloroplasztiszok DNS-polimerázát és aminoacil-tRNS-szintetázát kódolják. A riboszómális fehérjék többsége nukleáris gének ellenőrzése alatt áll. Mindezek az adatok arra késztetnek bennünket, hogy a kloroplasztiszokról, valamint a mitokondriumokról beszéljünk, mint korlátozott autonómiával rendelkező struktúrákról.
A fehérjék transzportja a citoplazmából a plasztidokba elvileg hasonlóan megy végbe, mint a mitokondriumokban. Itt azokon a helyeken, ahol a kloroplaszt külső és belső membránja összeér, csatornaképző integrált fehérjék találhatók, amelyek felismerik a citoplazmában szintetizált kloroplasztfehérjék szignálszekvenciáját, és a mátrix stromába szállítják. További szignálszekvenciák szerint a stromából importált fehérjék beépülhetnek plasztid membránokba (tilakoidok, stromalamellák, külső és belső membránok), vagy a stromában lokalizálhatók, riboszómák, a Calvin ciklus enzimkomplexei stb. részeként.
A szerkezet és az energiafolyamatok meglepő hasonlósága egyrészt a baktériumokban és mitokondriumokban, másrészt a kékalgákban és a kloroplasztiszokban erős érvként szolgál ezen organellumok szimbiotikus eredetének elmélete mellett. Ezen elmélet szerint az eukarióta sejt megjelenése a más sejtekkel való szimbiózis több szakaszán ment keresztül. Az első szakaszban az anaerob heterotróf baktériumok típusú sejtjei tartalmaztak aerob baktériumokat, amelyek mitokondriumokká alakultak. Ezzel párhuzamosan a gazdasejtben a prokarióta genofórból a citoplazmából izolált sejtmag alakul ki. Tehát heterotróf eukarióta sejtek keletkezhettek. Az elsődleges eukarióta sejtek és a kékalgák közötti ismétlődő endoszimbiotikus kapcsolatok olyan kloroplaszt típusú struktúrák megjelenéséhez vezettek bennük, amelyek lehetővé teszik a sejtek számára, hogy autoszintetikus folyamatokat hajtsanak végre, és nem függjenek a szerves szubsztrátok jelenlététől (236. ábra). Egy ilyen összetett élőrendszer kialakulása során a mitokondriumok és plasztidok genetikai információinak egy része megváltozhat, átkerülhet a sejtmagba. Így például a kloroplasztiszok 60 riboszómális fehérjéjének kétharmada a sejtmagban kódolódik, és a citoplazmában szintetizálódik, majd beépül a kloroplaszt riboszómákba, amelyek a prokarióta riboszómák összes tulajdonságával rendelkeznek. A prokarióta gének nagy részének ilyen átvitele a sejtmagba oda vezetett, hogy ezek a sejtszervecskék, megőrizve korábbi autonómiájuk egy részét, a sejtmag irányítása alá kerültek, amely nagyobb mértékben meghatározza az összes fő sejtfunkciót.
proplasztidok
Normál fényben a proplasztidok kloroplasztokká alakulnak. Először is nőnek, és a belső membránból hosszirányban elhelyezkedő membránredők képződnek. Némelyikük a plasztisz teljes hosszában kiterjed, és stromalamellákat képez; mások tilakoid lamellákat képeznek, amelyek felhalmozódnak és érett kloroplasztiszokból granákat képeznek. A plasztidok némileg eltérő fejlődése megy végbe a sötétben. Az etiolált palántákban kezdetben a plasztidok, etioplasztok térfogatának növekedése következik be, de a belső membránok rendszere nem lamellás struktúrákat épít, hanem kis buborékok tömegét képezi, amelyek külön zónákban halmozódnak fel, és akár összetett rácsszerkezeteket is kialakíthatnak. (prolamelláris testek). Az etioplasztok membránjai protoklorofillt, a sárga klorofill előfutárát tartalmazzák. A fény hatására etioplasztokból kloroplasztok képződnek, a protoklorofill klorofilllá alakul, új membránok, fotoszintetikus enzimek és az elektrontranszport lánc komponensei szintetizálódnak.
A sejtek megvilágításakor a membrán vezikulák és tubulusok gyorsan átrendeződnek, amiből a lamellák és tilakoidok teljes rendszere fejlődik ki, ami a normál kloroplasztiszra jellemző.
A leukoplasztok fejlett lamelláris rendszer hiányában különböznek a kloroplasztiszoktól (226. b ábra). A tárolószövetek sejtjeiben találhatók. Bizonytalan morfológiájuk miatt a leukoplasztokat nehéz megkülönböztetni a proplasztidoktól és néha a mitokondriumoktól. A proplasztidokhoz hasonlóan lamellában szegények, de fény hatására normális tilakoid struktúrákat képesek kialakítani és zöld színt kapni. Sötétben a leukoplasztok különféle tartalék anyagokat halmozhatnak fel a prolamelláris testekben, és másodlagos keményítőszemcsék rakódnak le a leukoplasztok strómájában. Ha a kloroplasztiszokban úgynevezett tranziens keményítő rakódik le, ami itt csak a CO2 asszimilációja során van jelen, akkor a keményítő valódi raktározása a leukoplasztokban valósulhat meg. Egyes szövetekben (gabona endospermiumban, rizómákban és gumókban) a leukoplasztokban lévő keményítő felhalmozódása a plasztisz stromában elhelyezkedő raktározó keményítőszemcsékkel teljesen megtelt amiloplasztok képződéséhez vezet (226c. ábra).
A magasabb rendű növények plasztiszainak másik formája a kromoplaszt, amely általában a karotinoidok felhalmozódása következtében sárgul (226d. ábra). A kromoplasztok kloroplasztiszokból és sokkal ritkábban leukoplasztjaikból képződnek (például a sárgarépa gyökerében). A kloroplasztiszok elszíneződésének és változásának folyamata könnyen megfigyelhető a szirmok fejlődése során vagy a termések beérésekor. Ugyanakkor a plasztidokban sárga színű cseppek (gömböcskék) halmozódhatnak fel, vagy kristály formájú testek jelennek meg bennük. Ezek a folyamatok a plasztidban lévő membránok számának fokozatos csökkenésével, a klorofill és a keményítő eltűnésével járnak. A színes gömböcskék képződésének folyamatát az magyarázza, hogy a kloroplasztiszok lamelláinak elpusztulásakor lipidcseppek szabadulnak fel, amelyekben a különböző pigmentek (például karotinoidok) jól oldódnak. Így a kromoplasztok a plasztidok degeneráló formái, amelyek lipofanerózisnak, a lipoprotein komplexek lebomlásának vannak kitéve.
A sejt egy összetett szerkezet, amely sok komponensből, úgynevezett organellumokból áll. Ugyanakkor a kompozíció növényi sejt némileg különbözik az állattól, és a fő különbség a jelenlétben rejlik plasztidok.
Kapcsolatban áll
A sejtelemek leírása
A sejtek mely összetevőit nevezzük plasztidoknak. Ezek összetett szerkezetű és funkciójú szerkezeti sejtszervecskék, amelyek fontosak a növényi szervezetek életében.
Fontos! A plasztidok proplasztidokból képződnek, amelyek a merisztéma vagy oktatási sejtek belsejében helyezkednek el, és sokkal kisebbek, mint egy érett organoid. És ezek is, mint a baktériumok, kétfelé oszlanak szűkítéssel.
Mi van plasztidok szerkezet mikroszkóp alatt nehezen láthatóak, a sűrű héj miatt nem áttetszőek.
A tudósoknak azonban sikerült kideríteniük, hogy ennek az organoidnak két membránja van, belsejében stróma van, amely a citoplazmához hasonló folyadék.
A belső membrán redői halomba rakva grana-t alkotnak, amely összekapcsolható.
Benne is vannak riboszómák, lipidcseppek, keményítőszemcsék. Még a plasztidoknak, különösen a kloroplasztiszoknak is megvannak a maguk molekulái.
Osztályozás
Színük és funkcióik szerint három csoportra oszthatók:
- kloroplasztiszok,
- kromoplasztok,
- leukoplasztok.
Kloroplasztok
A legmélyebben tanulmányozottak zöld színűek. A növények leveleiben, néha szárában, gyümölcsében, sőt gyökereiben is megtalálható. Kinézetre úgy néznek ki, mint a 4-10 mikrométer méretű lekerekített szemcsék. Kis méret és nagy mennyiség nagymértékben megnöveli a munkafelület területét.
Színük eltérő lehet, ez a bennük lévő pigment típusától és koncentrációjától függ. Alapvető pigment - klorofill, xantofill és karotin is jelen van. A természetben 4 fajta klorofill van, amelyeket latin betűkkel jelölnek: a, b, c, e. Az első két típus magasabbrendű növények sejtjeit és zöld algákat tartalmaz, a kovamoszatoknak csak fajtái vannak - a és c.
Figyelem! Más organellumokhoz hasonlóan a kloroplasztiszok is képesek öregedni és lebomlani. A fiatal struktúra megosztottságra és aktív munkára képes. Idővel szemcséik elpusztulnak, és a klorofill lebomlik.
A kloroplasztiszok fontos funkciót töltenek be: bennük a fotoszintézis folyamata megy végbe- a napfény átalakítása szénhidrátképző kémiai kötések energiájává. Ugyanakkor együtt mozoghatnak a citoplazma áramával, vagy aktívan mozoghatnak önállóan. Tehát gyenge megvilágítás mellett nagy mennyiségű fénnyel halmozódnak fel a cella falai közelében, és nagyobb területtel fordulnak felé, és nagyon aktív fényben éppen ellenkezőleg, élesen állnak.
Kromoplasztok
 Helyettesítik a megsemmisült kloroplasztiszokat, sárga, piros és narancssárga árnyalatúak. A színezék a karotinoidok miatt képződik.
Helyettesítik a megsemmisült kloroplasztiszokat, sárga, piros és narancssárga árnyalatúak. A színezék a karotinoidok miatt képződik.
Ezek az organellumok a növények leveleiben, virágaiban és gyümölcseiben találhatók. A forma lehet kerek, téglalap vagy akár tű alakú is. Szerkezete hasonló a kloroplasztiszokhoz.
Fő funkció - színezés virágok és gyümölcsök, ami vonzza a beporzó rovarokat és az állatokat, amelyek megeszik a gyümölcsöket, és így hozzájárulnak a növény magjainak szétszóródásához.
Fontos! A tudósok spekulálnak a szerepről kromoplasztok a sejt redox folyamataiban fényszűrőként. Figyelembe veszik a növények növekedésére és szaporodására gyakorolt hatásukat.
Leukoplasztok
Adat a plasztidoknak van különbségek a szerkezete és funkciója. A fő feladat a tápanyagok jövőre való tárolása, így azok elsősorban a termésekben, de előfordulhatnak a növény megvastagodott és húsos részeiben is:
- gumók
- rizómák,
- gyökérnövények,
- izzók és mások.
 színtelen színezés nem teszi lehetővé azonosításukat. a sejt szerkezetében viszont jól láthatóak a leukoplasztok, ha kis mennyiségű jódot adunk hozzá, ami a keményítővel kölcsönhatásba lépve kékre festi őket.
színtelen színezés nem teszi lehetővé azonosításukat. a sejt szerkezetében viszont jól láthatóak a leukoplasztok, ha kis mennyiségű jódot adunk hozzá, ami a keményítővel kölcsönhatásba lépve kékre festi őket.
A forma kerekhez közeli, míg belül a membránrendszer gyengén fejlett. A membránredők hiánya segíti az organoidot az anyagok tárolásában.
A keményítőszemcsék megnövekednek, és könnyen elpusztítják a plasztid belső membránjait, mintha megfeszítenék. Ez lehetővé teszi több szénhidrát tárolását.
Más plasztidokkal ellentétben ezek formalizált formában tartalmaznak egy DNS-molekulát. Ugyanakkor a klorofill felhalmozásával, a leukoplasztok kloroplasztokká alakulhatnak.
A leukoplasztok funkciójának meghatározásakor meg kell jegyezni a specializációjukat, mivel több típus létezik, amelyek bizonyos típusú szerves anyagokat tárolnak:
- az amiloplasztok keményítőt halmoznak fel;
- az oleoplasztok zsírokat termelnek és tárolnak, míg az utóbbiak a sejtek más részein raktározódnak;
- a proteinoplasztok "védik" a fehérjéket.
 A felhalmozódáson kívül anyaghasító funkciót is elláthatnak, amihez olyan enzimek kapcsolódnak, amelyek energia- vagy építőanyaghiány esetén aktiválódnak.
A felhalmozódáson kívül anyaghasító funkciót is elláthatnak, amihez olyan enzimek kapcsolódnak, amelyek energia- vagy építőanyaghiány esetén aktiválódnak.
Ilyen helyzetben az enzimek elkezdik a raktározott zsírokat és szénhidrátokat monomerekre bontani, hogy a sejt megkapja a szükséges energiát.
A plasztidok minden fajtája, annak ellenére szerkezeti jellemzők képesek legyenek átváltozni egymásba. Tehát a leukoplasztok kloroplasztokká alakíthatók, ezt a folyamatot akkor látjuk, amikor a burgonyagumó zöldre vált.
Ugyanakkor ősszel a kloroplasztiszok kromoplasztokká alakulnak, aminek következtében a levelek sárgulnak. Minden sejt csak egyféle plasztidot tartalmaz.
Eredet
Számos eredetelmélet létezik, ezek közül kettő a legésszerűbb:
- szimbiózis,
- abszorpció.
Az első a sejtképződést több lépésben lezajló szimbiózis folyamatnak tekinti. Lefolyása során a heterotróf és autotróf baktériumok egyesülnek, kölcsönös előnyök megszerzése.
A második elmélet a sejt kialakulását a kisebb szervezetek nagyobb szervezetek általi abszorpciója révén vizsgálja. Ebben az esetben azonban nem emésztődnek meg, beépülnek a baktérium szerkezetébe, abban látják el funkciójukat. Ez a szerkezet kényelmesnek bizonyult, és előnyt biztosított az organizmusoknak másokkal szemben.
Plasztidok típusai növényi sejtben
Plasztidok - funkcióik a sejtben és típusai
Következtetés
A növényi sejtekben található plasztidok egyfajta „gyár”, ahol a termelés mérgező intermedierekkel, nagy energiájú és szabad gyökös átalakulási folyamatokkal társul.
A plasztidok (görögül plasztidok – létrejövő, képződő) fotoszintetikus eukarióta organellumok membránszervecskéi – magasabbrendű növények, alacsonyabb algák, egyes egysejtűek. A plasztidok minden típusú növényi sejtben jelen vannak, mindegyik típusnak megvan a maga saját készlete ezekből az organellumokból. Minden plasztidnak számos közös jellemzője van. Saját genetikai berendezéssel rendelkeznek, és két koncentrikus membránból álló héj veszi körül.
Minden plasztid proplasztidokból fejlődik ki. A merisztémasejtekben jelenlévő kisméretű organellumok, amelyek sorsát a differenciált sejtek szükségletei határozzák meg. A plasztidok minden típusa egyetlen genetikai sorozatot képvisel.
A leukoplasztok (görögül leucos - fehér) színtelen plasztidok, amelyek a növényi szervek sejtjeiben találhatók, színtelen. Lekerekített képződmények, amelyek legnagyobb mérete 2-4 mikron. Két membránból álló membrán veszi körül őket, amelyek belsejében fehérje stroma található. A leukoplasztok stromája kisszámú hólyagot és lapos ciszternát - lamellákat - tartalmaz. A leukoplasztok képesek kloroplasztokká fejlődni, fejlődésük folyamata a méret növekedésével, a belső szerkezet komplikációjával és a zöld pigment - klorofill képződésével jár. A plasztidok ilyen átrendeződése például a burgonyagumók zöldítése során következik be. A leukoplasztok is képesek kromoplasztokká átalakulni. Egyes szövetekben, például a gabonaszemek endospermiumában, a rizómákban és gumókban, a leukoplasztok a keményítő tartalék raktárává - amiloplasztokká - alakulnak. Az egyik formából a másikba való ontogenetikai átmenetek visszafordíthatatlanok, a kromoplaszt nem képezhet sem kloroplasztot, sem leukoplasztot. Hasonlóképpen, a kloroplaszt nem tud visszatérni a leukoplaszt állapotába.
A kloroplasztok (klórzöld) a plasztidok fő formája, amelyekben a fotoszintézis végbemegy. A magasabb rendű növények kloroplasztjai lencse alakú képződmények, amelyek szélessége a rövid tengely mentén 2-4 µm, a hosszú tengely mentén 5 µm vagy több. A különböző növények sejtjeiben a kloroplasztiszok száma nagyon eltérő, a magasabb rendű növények sejtjei 10-30 kloroplasztot tartalmaznak. Körülbelül ezret találtak belőlük a bozontos palánkszövet óriás sejtjeiben. Az algákban található kloroplasztokat eredetileg kromatoforoknak hívták. A zöld algák sejtenként egy kromatoforral rendelkezhetnek, az euglenoidokban és a dinoflagellatákban a fiatal sejtek 50-80 kloroplasztot tartalmaznak, az öregek - 200-300. Az algák kloroplasztiszai lehetnek csésze alakúak, szalag alakúak, spirálisak, lamellásak, csillagszerűek, szükségszerűen tartalmaznak sűrű fehérjetermészetű képződményt - pirenoidokat, amelyek körül a keményítő koncentrálódik.
A kloroplasztiszok ultrastruktúrája nagy hasonlóságot mutat a mitokondriumokkal, elsősorban a kloroplasztisz membrán - perisztromium - szerkezetében. Két membrán veszi körül, amelyeket egy keskeny, körülbelül 20-30 nm széles membránköz választ el. A külső membrán nagy áteresztőképességű, míg a belső kevésbé áteresztő, és speciális transzportfehérjéket hordoz. Hangsúlyozni kell, hogy a külső membrán nem átjárható az ATP számára. A belső membrán egy nagy központi régiót vesz körül - a stromát, amely a mitokondriális mátrix analógja. A kloroplaszt sztróma különféle enzimeket, riboszómákat, DNS-t és RNS-t tartalmaz. Jelentős különbségek is vannak. A kloroplasztiszok sokkal nagyobbak, mint a mitokondriumok. Belső membránjuk nem képez krisztákat és nem tartalmaz elektrontranszport láncot. A kloroplaszt összes legfontosabb funkcionális eleme a harmadik membránban található, amely lapított korong alakú tasakok - tilakoidok - csoportjait alkotja; ezt tilakoid membránnak nevezik. Ez a membrán összetételében pigment-fehérje komplexeket, elsősorban klorofillt, a karotinoidok csoportjába tartozó pigmenteket tartalmaz, amelyek közül gyakori a karotin és a xantofil. Ezenkívül a tilakoid membránban elektrontranszport láncok komponensei is megtalálhatók. A tilakoidok belső üregei hozzák létre a kloroplaszt harmadik belső kompartmentjét, a tilakoid teret. A tilakoidok halmokat alkotnak – néhány darabtól 50 vagy több darabig terjedő gránát tartalmaznak. A szemcsék mérete a bennük lévő tilakoidok számától függően elérheti a 0,5 μm-t is, ilyenkor fénymikroszkóppal is megfigyelhetőek. A gránában lévő tilakoidok szorosan kapcsolódnak egymáshoz, membránjuk érintkezési pontján a rétegvastagság körülbelül 2 nm. A Grana a tilakoidokon kívül stromalamellák szakaszait is tartalmazza. Ezek lapos, hosszúkás, perforált tasakok, amelyek a kloroplasztisz párhuzamos síkjaiban helyezkednek el. Nem metszik egymást és zárva vannak. A stroma lamellák egyes gránákat kötnek össze. Ugyanakkor a tilakoid üregek és a lamellák üregei nem kapcsolódnak egymáshoz.
A kloroplasztiszok feladata a fotoszintézis, a napfény energiája hatására szerves anyagok képződése szén-dioxidból és vízből. Ez az egyik legfontosabb biológiai folyamat, amely folyamatosan és hatalmas léptékben zajlik bolygónkon. A földgömb növényzete minden évben több mint 100 milliárd tonna szerves anyagot képez, mintegy 200 milliárd tonna szén-dioxidot asszimilálva, és mintegy 145 milliárd tonna szabad oxigént bocsát ki a környezetbe.
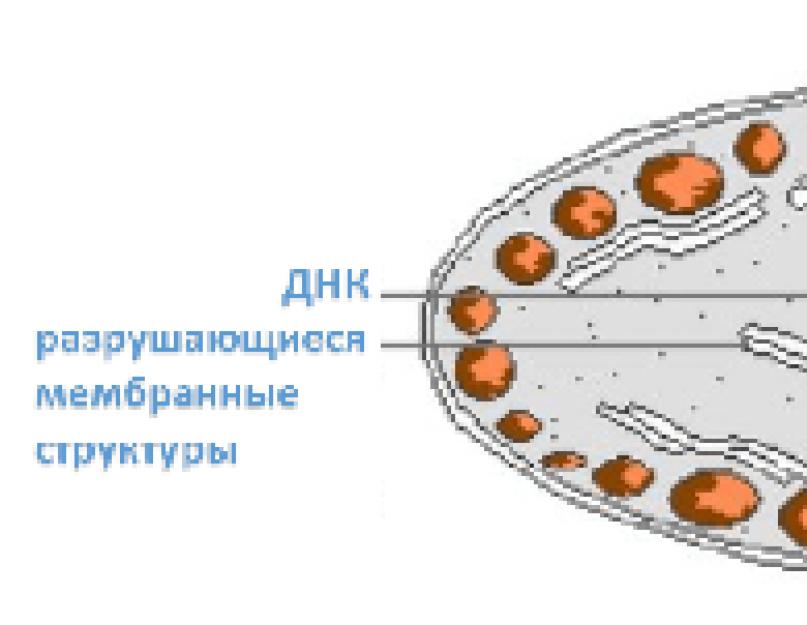
Kromoplasztok Ezek egy növényi sejt plasztidjai, amelyek sárga-narancs színűek. Szenilis, lebontó sejtszervecskékként definiálhatók, a kloroplasztiszok pusztulása során keletkeznek. Ezt bizonyítja a plasztidok kémiai összetétele. Ha a kloroplasztiszokban a fehérjék a teljes tömegük körülbelül 50%-át, a lipidek 30%-át teszik ki, akkor a kromoplasztokban ez az arány a következőképpen változik: 22% fehérjék, 58% lipidek, DNS már nem mutatható ki. A kromoplasztok színe a karotinoidok jelenlététől és a klorofill pusztulásától függ. A klorofill lebontásából származó nitrogéntartalmú vegyületek (pirrolszármazékok) ugyanúgy áramlanak ki a levelekből, mint a membránok fehérje-lipid rendszerének lebomlása során keletkező fehérjék. A lipidek a perisztromiumban maradnak. A karotinoidok feloldódnak bennük, sárga és narancssárga tónusúvá festve a plasztidokat. A kromoplasztok képződése kloroplasztiszokból kétféleképpen történik. Például a boglárkában a kromoplasztok halványzöld keményítőtartalmú kloroplasztiszokból képződnek. A klorofill és a keményítő fokozatosan eltűnik, növekszik a sárga pigment tartalma, amely lipidcseppekben oldódik, gömböcskéket képezve. A gömböcskék képződésével egyidejűleg a kloroplaszt lamellás szerkezetének végső megsemmisülése következik be. A kialakult kromoplasztban csak a perisztromium őrződik meg, teljes belső felületét golyócskák borítják, a plasztisz közepe optikailag üresnek tűnik. A kromoplasztok szerepe a sejtben nem tisztázott. De a növényi szervezet egésze számára ezek a plasztidok fontos szerepet játszanak, mivel a növény azon szervei, amelyekben a fotoszintézis leáll, vonzóvá válnak a rovarok, madarak és más növényeket beporzó, gyümölcseiket és magjaikat terjesztő állatok számára. A levelek őszi sárgulása során a kloroplasztiszok pusztulása és a kromoplasztok képződése a fehérjék és nitrogéntartalmú vegyületek hasznosulásához vezet, amelyek a növény más szerveibe áramlanak még a lombhullás előtt.



